Abstract
Background
Marinobufagenin (MBG) promotes natriuresis via inhibition of renotubular Na/K-ATPase (NKA) and causes vasoconstriction via inhibition of vascular NKA. Atrial natriuretic peptide (ANP), via cyclic guanosine monophosphate (cGMP)/protein kinase G (PKG)-dependent mechanism, sensitizes renal NKA to MBG but reduces MBG-induced inhibition of vascular NKA. As aging is associated with a downregulation of cGMP/PKG signaling, we hypothesized that in older rats, ANP would not potentiate renal effects of MBG and would not oppose vascular effects of MBG.
Methods
In younger (3-month-old) and older (12-month old) Sprague–Dawley rats, we compared SBP, natriuresis, activity of NKA in aorta and renal medulla, and levels of MBG and α-ANP at baseline and following acute NaCl loading (20%, 2.5 ml/kg, intraperitoneally), and studied modulation of MBG-induced NKA inhibition by α-ANP in vitro.
Results
As compared with younger rats, NaCl-loaded older rats exhibited a greater MBG response, greater SBP elevation (25 vs. 10 mmHg, P<0.01) and greater inhibition of NKA in aorta (39 vs. 7%, P<0.01), 30% less natriuresis, and less inhibition of renal NKA (25 vs. 42%, P<0.05) in the presence of comparable responses of α-ANP and cGMP. In aorta and kidney of older rats, the levels of PKG were reduced, the levels of phosphodiesterase-5 were increased compared with that in young rats, and α-ANP failed to modulate MBG-induced NKA inhibition.
Conclusion
Age-associated downregulation of cGMP/PKG-dependent signaling impairs the ability of ANP to modulate the effects of MBG on the sodium pump, which contributes to salt sensitivity.
Keywords: aging, cyclic guanosine monophosphate, dietary sodium, hypertension, marinobufagenin, Na/K-ATPase, natriuretic peptides, protein kinase G, salt sensitivity
INTRODUCTION
A shift in the balance between pressor and natriuretic responses to dietary NaCl intake is one of the factors that underlie age-associated increase in the salt sensitivity of blood pressure [1–3]. Numerous factors are implicated in the regulation of sodium reabsorption in the kidney, including digitalis-like cardiotonic steroids (CTSs) [4] and atrial natriuretic peptide (ANP) [5,6]. CTSs promote natriuresis via inhibition of the Na/K-ATPase (NKA) in the proximal tubules and thick ascending limb (TAL) of Henle’s loop, but excessive CTS production induces vasoconstriction leading to hypertension [8]. ANP exhibits natriuretic effect via activation of cyclic nucleotide-gated cation channels in the inner medullary collecting duct and via modulation of NKA activity by cyclic guanosine monophosphate (cGMP) and protein kinase G (PKG) in the TAL of Henle’s loop [5,7]. Because PKG is implicated in regulatory phosphorylation of the NKA, a receptor for CTS [9], and because in TAL, ANP acts via PKG-dependent mechanism [5], this segment of nephron represents a site for interaction between CTS and ANP. Thus, ANP, acting via cGMP/PKG-dependent mechanism, was shown to sensitize NKA to a bufadienolide CTS marinobufagenin (MBG) in a suspension of outer medullary tubules, in which fragments of TAL comprise 90% of tissue mass [10]. Although only about 15–25% of filtered sodium is reabsorbed in TAL, the importance of this segment of nephron in sodium metabolism could be illustrated by the fact that a reduction in salt reabsorption by TAL underlies pathogenesis of Bartter’s disorders of diverse genetic origins [11].
Previously, we demonstrated that the plasma levels of α-hANP and of MBG covary in hypertensive patients with heart failure [12]. Because ANP acts as a natriuretic and a vasorelaxant [5], and MBG elicits natriuresis and induces vasoconstriction [4,13], we hypothesized that simultaneous production of these two hormones under the conditions of salt loading and volume expansion makes teleological sense; that is, although both hormones exhibit synergism with respect to renal sodium excretion, ANP could potentially offset the excessive vasoconstriction caused by MBG. In support of this hypothesis, we demonstrated that low concentrations of ANP, acting via cGMP-dependent mechanism, sensitize NKA from rat renal medulla to MBG, but reduce MBG sensitivity of the NKA from vascular sarcolemma [10]. Later, we compared patterns of responses of blood pressure, renal sodium excretion, MBG, ANP, cGMP, and renal and vascular sodium pump activity to acute NaCl loading in Dahl salt-sensitive rats (Dahl-S) and in relatively salt-insensitive Sprague–Dawley rats [14]. We found that in response to NaCl loading, Dahl-S exhibited a greater pressor response and a greater inhibition of NKA in vascular sarcolemma in the presence of a lesser natriuretic response and a lesser inhibition of renotubular NKA, as compared with that in Sprague–Dawley rats [14]. The response of MBG to NaCl loading was comparable in both strains, but ANP response to NaCl was blunted in Dahl-S [14], suggesting that in salt-sensitive rats, a decline in ANP–cGMP–PKG signaling reduces the sensitivity of the renal sodium pump to the natriuretic effect of MBG but sensitizes vascular NKA to the pressor action of this hormone.
As aging is associated with both increased salt sensitivity of blood pressure [1,2] and decline in cGMP/PKG-dependent signaling [15–17], we hypothesized that the above scenario described in Dahl-S rats may also be relevant to the development of age-dependent salt sensitivity in a normotensive rat. In the present study, we compared the effect of acute NaCl loading on MBG, ANP, renal and vascular NKA, blood pressure, and renal sodium excretion in younger (3-month-old) and older (12-month-old) Sprague–Dawley rats.
METHODS
Experimental design
The experimental protocol was approved by the Animal Care and Use Committee of the National Institute on Aging. Twenty-four 3-month-old (321±7 g) and 24, 12-month-old (561±12 g) male Sprague–Dawley rats (Charles River Laboratories, Wilmington, Massachusetts, USA) were studied after 1 week of adaptation to the laboratory environment, metabolic chambers, and measurement of blood pressure via tail-cuff plethysmography (IITC Inc., Life Science, Woodland Hills, California, USA). Following baseline urine collection and blood pressure measurement, 12 younger and 12 older rats were sacrificed (below) serving as controls, and a single injection of 2.5 ml/kg of 20% NaCl was intraperitoneally administered to 12 rats of each age group under light anesthesia (25 mg/kg Ketaset; Fort Dodge Animal Health, Fort Dodge, Iowa, USA) [14]. Then, the animals were placed in metabolic chambers and urine was collected hourly for determination of renal excretion of sodium, MBG, and cGMP. Within 2 h following acute NaCl loading, SBP was measured and the animals were anesthetized with 60 mg/kg Ketaset and exsanguinated from the abdominal aorta. Plasma was collected for determination of MBG and α-ANP-1–28. Kidneys and aortae were collected for measurement of NKA activity (below).
The concentration of sodium in the urine was measured using Roche-Hitachi 917 (Roche, Vienna, Austria), and urinary creatinine was measured using creatinine urinary assay kit (Cayman Chemical, Ann Arbor, Michigan, USA). Plasma electrolytes, plasma creatinine, and plasma glucose were measured by i-Stat analyzer (Abbott Laboratories, Abbott Park, Illinois, USA). Fractional excretion of sodium (FeNa) was calculated as FENa = uNa × pCr × 100/(pNa × uCr)), where uNa and pNa are urine and plasma sodium concentrations (mmol/l), respectively, and uCr and pCr are urine and plasma creatinine concentrations (mg/dl), respectively.
Preparation of vascular and renal membranes
Membranes from aortae were prepared as described previously [10,14]. Aortic rings were minced by scissors, processed with Polytron 20S homogenizer (Kinematica, Lucerne, Switzerland) in 250mmol/l sucrose and 5 mmol/l histidine solution (4°C, pH 7.4), and centrifuged (6000 g for 15 min at 4°C) in a Beckman L8-N centrifuge. The pellet was rehomogenized and centrifuged (6000g for 15 min at 4°C). The supernatant was respun at 10 000g for 30 min at 4°C and the resultant supernatant was centrifuged (Beckman L8-N, 148 000g for 90 min at 4°C). The pellet was suspended in a homogenizing medium to a protein concentration of 1mg/ml and stored in liquid nitrogen. Membranes from renal outer medulla (suspension of outer medullary tubules, in which fragments of the TAL comprise 90% of tissue mass) were prepared as described previously [10,14]. Medulla slices were homogenized in a solution containing (in mmol/l) sucrose 250, histidine 30, imidazole 5, EDTA 1 (4°C, pH 7.4), then centrifuged (6000g for 15 min at 4°C) in a Beckman L8-N centrifuge. The supernatant was respun at 15 000g for 30 min at 4°C and the resultant supernatant centrifuged (Beckman L8-N, 148 000g for 90 min at 4°C). The pellet was re-suspended in a homogenizing medium to a protein concentration of 4 mg/ml and stored in liquid nitrogen. NKA activity was measured as reported previously [10,14]. To increase the permeability of membranes vesicles, membranes were pretreated with alamethicin (0.5 mg/mg protein). Aliquots of membranes from aortic sarcolemma or renal medulla (0.5 mg protein/100ml) were preincubated for 60 min at 37°C with compounds as described and then incubated for 1 h (for aorta) or 15 min (for renal medulla) at 37°C in 96-well low-binding polystyrene plates in a medium containing NaCl 100 mmol/l, KCl 4 mmol/l, MgCl2 3 mmol/l, EDTA 1 mmol/l, Tris 50 mmol/l, ATP 3 mmol/l, and NaN3 5 mmol/l, pH was 7.4. The reaction was stopped by quenching solution containing 1.25N H2SO4. NKA activity was estimated as the difference between total ATPase activity in the presence and absence of 1 mmol/l ouabain.
Na/K-ATPase phosphorylation
NKA phosphorylation was assessed as previously described [6]. Membranes from renal medulla and aortic sarcolemma were pretreated with alamethicin (0.5 mg/mg protein) and preincubated in the presence of compounds studied for 30 min at 32°C in a buffer containing Tris phosphate 10 mmol/l, magnesium acetate 5 mmol/l, and CaCl2 0.5 mmol/l. As basal levels of NKA phosphorylation in aortic sarcolemma were very low, phosphorylation was enhanced by preincubation of membranes with 50 nmol/l phorbol 12,13-diacetate for 3 min before the addition of the studied compounds, as previously reported [18]. Phosphorylation was initiated by the addition of [g-32P]ATP (1000 cpm/pmol) to a final concentration of 70 mmol/l. The reaction was quenched with an equal volume of Novex Tri-Glycine SDS sample buffer. Electrophoresis was performed as described below (western blotting). Phosphoproteins were visualized by a 12–24-h exposure of nitrocellulose membranes on Kodak XAR-5 film at 4°C. The optical density of bands, corresponding to NKA (112 kDa), was quantified via laser densiometry using Bio-Rad Gel Doc 1000 software (Bio-Rad Laboratories, Berkeley, California, USA). To verify the localization of α-1 NKA, the same nitrocellulose membranes were further exposed to the mouse monoclonal a α-1 NKA antibody (clone 464.4; Upstate Biotechnologies, Lake Placid, Massachusetts, USA, 1 : 2000) (western blotting, below).
Western blotting
Solubilized proteins from aortic sarcolemma and renal medulla were separated by 10% Tris-glycine polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The proteins were visualized using monoclonal mouse anti α1-NKA (Upstate Biotechnologies; 1 : 2000), polyclonal rabbit anti-α2-NKA (Upstate Biotechnologies; 1 : 500), monoclonal mouse anti-α3-NKA (Affinity Bio-Reagents Inc., Golden, Colorado, USA; 1 : 500) antibody, polyclonal rabbit anti-PKG I (cGPK1b) antibody (Stressgen, Victoria, British Columbia, Canada; 1 : 2000), polyclonal goat anti-PKGII (cGKII) antibody (Santa Cruz Biotechnology Inc., Santa Cruz, California, USA; 1 : 2500), and polyclonal rabbit anti-PDE-5A antibody (Santa Cruz Biotechnology Inc.; 1 : 1000) followed by incubation with peroxidase-conjugated antimouse, antirabbit antiserum (AmershamCorp., Arlington Heights, Illinois, USA; 1 : 1000), or antigoat antiserum (Santa Cruz Biotechnology Inc.; 1 : 1000). Bands were visualized by 1–5-min exposure of nitrocellulose membrane on Kodak SAR5 film and optical density was quantified by laser densiometry. To normalize the levels of proteins against the levels of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), membranes were stripped and reprobed with a rabbit monoclonal anti-GAPDH antibody (Cell Signaling Technology, Danvers, Massachusetts, USA).
Immunoassays
Plasma and urine samples were extracted on Sep-Pak C-18 cartridges (Waters, Milford, Massachusetts, USA) and MBG competitive fluoroimmunoassay based on a monoclonal murine anti-MBG 4G4 antibody was performed as described recently in detail [19]. The cross-reactivity of 4G4 anti-MBG antibody is as follows: MBG 100%, ouabain 0.005%, digoxin 0.03%, digitoxin less than 0.001%, bufalin 0.08%, cinobufagin 0.07%, cinobufatalin 40%, prednisone less than 0.001%, spironolactone less than 0.001%, proscillaridin less than 0.001%, and progesterone less than 0.001%. Plasma levels of rat ANP 1–28 were measured using enzyme immunoassay kit (Peninsula Laboratories, San Carlos, California, USA). Urinary cGMP was measured using enzyme immunoassay kit (Cayman Chemical).
Statistics
The results are reported as mean±SEM. The significance of differences among the measured variables was assessed by one-way analysis of variance, followed by Newman–Keuls test, and by two-way analysis of variance, where appropriate (GraphPad Prism software; GraphPad Inc., San Diego, California, USA). A P value less than 0.05 was considered statistically significant.
RESULTS
Data on the effects of NaCl loading on blood pressure, FeNa, diuresis, plasma sodium and potassium, and NKA activity are summarized in Fig. 1 and Table 1. As compared with 3-month-old rats, older animals in response to NaCl loading exhibited a greater pressor response and a reduced natriuretic response (Fig. 1) in the presence of comparable urine excretion at baseline and following NaCl loading (Table 1).
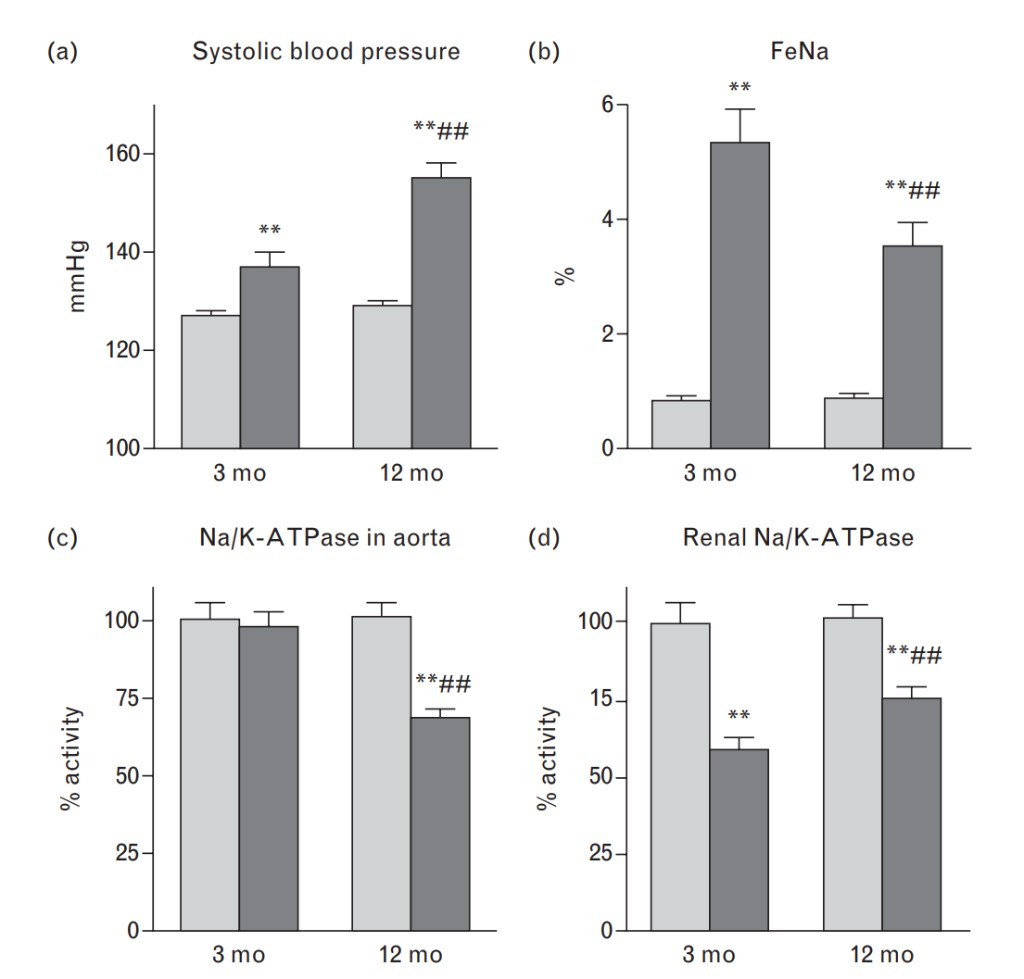
Table 1
Urine excretion, plasma electrolytes and hematocrit
| 3-month-old | 12-month-old | |||
|---|---|---|---|---|
| Baseline | NaCl | Baseline | NaCl | |
| Urine excretion (mL/hr) | 2.2 ± 0.3 | 7.4 ± 0.8* | 2.0 ± 0.3 | 6.9 ± 1.0* |
| Plasma Na (mmol/L) | 143 ± 1 | 145 ± 1* | 144 ± 1 | 149 ± 1*# |
| Plasma K (mmol/L) | 4.2 ± 0.1 | 4.0 ± 0.1 | 4.1 ± 0.1 | 4.4 ± 0.1 |
| Hematocrit (%) | 41 ± 1 | 43 ± 0.4* | 42 ± 1 | 45 ± 1* |
| Plasma glucose (mg/dl) | 291 ± 8 | 306 ± 5 | 313 ± 13 | 309 ± 15 |
At baseline, the activity of NKA in older rats was lower than that in 3-month-old animals in both aortic sarcolemma (4.1±0.1 vs. 5.1±0.1mmol Pi/mg protein/h, respectively; P<0.01) and renal outer medulla (28.9±1.4 vs. 34.8±1.6mmol Pi/mg protein/h, respectively; P<0.02). NaCl loading was associated with a 7% inhibition of NKA in aortic sarcolemma of younger animals, but produced a 39% NKA inhibition in 12-month-old rats (Fig. 1c). The pattern of responses of renal NKA to NaCl loading was opposite to that observed in aorta, and whereas NaCl loading induced 42% inhibition of renal NKA in 3-monthold rats, NKA was inhibited by only 25% in older animals (Fig. 1d).
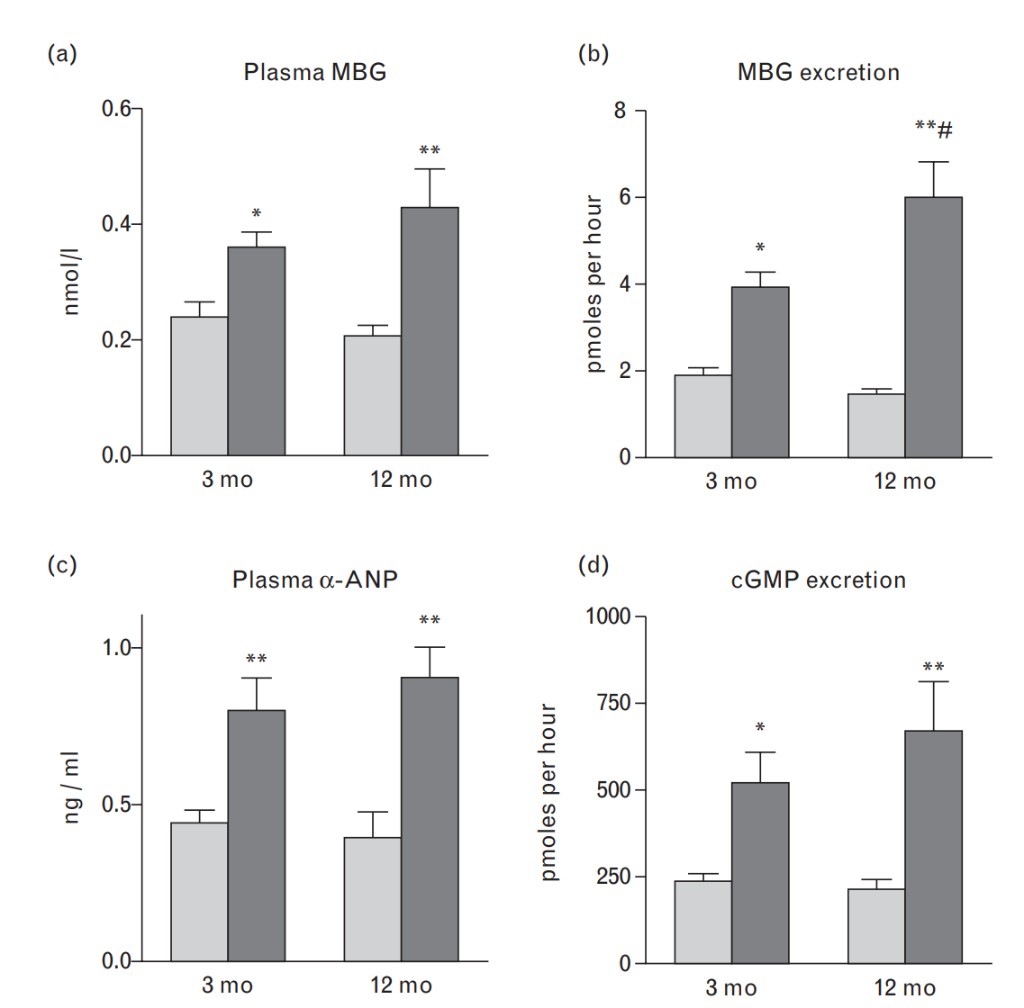
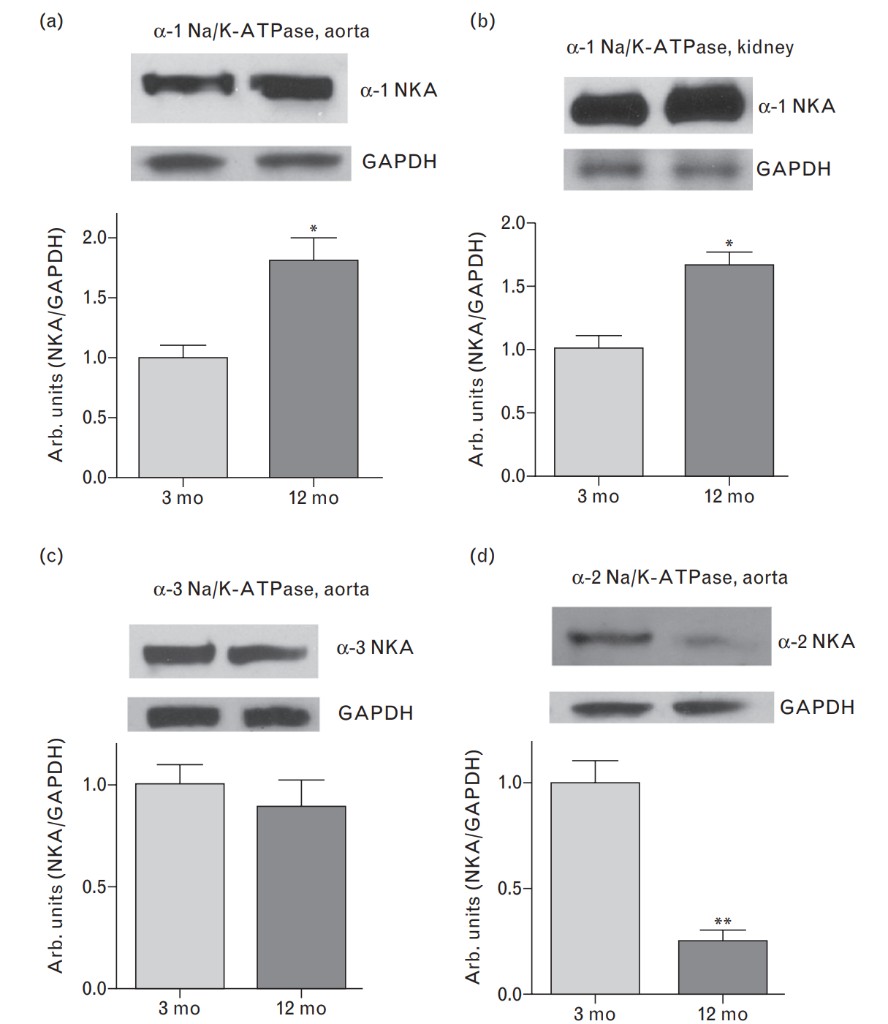
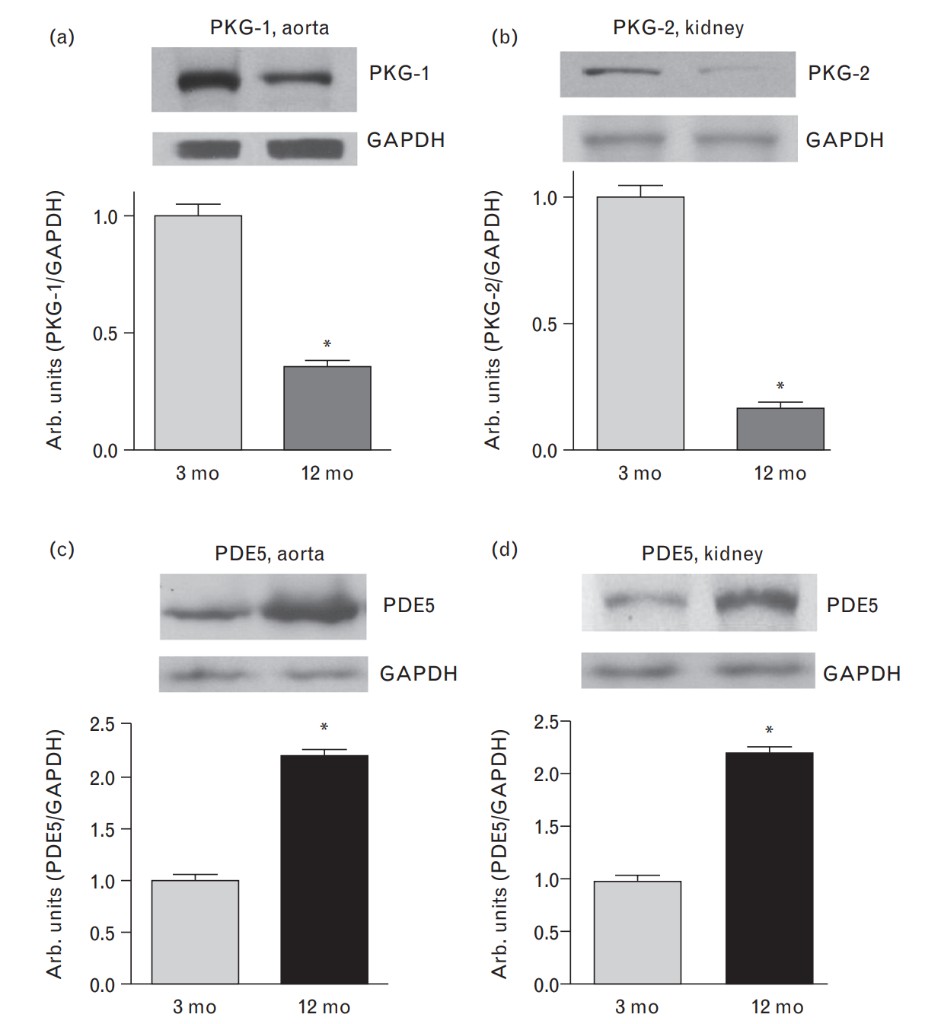
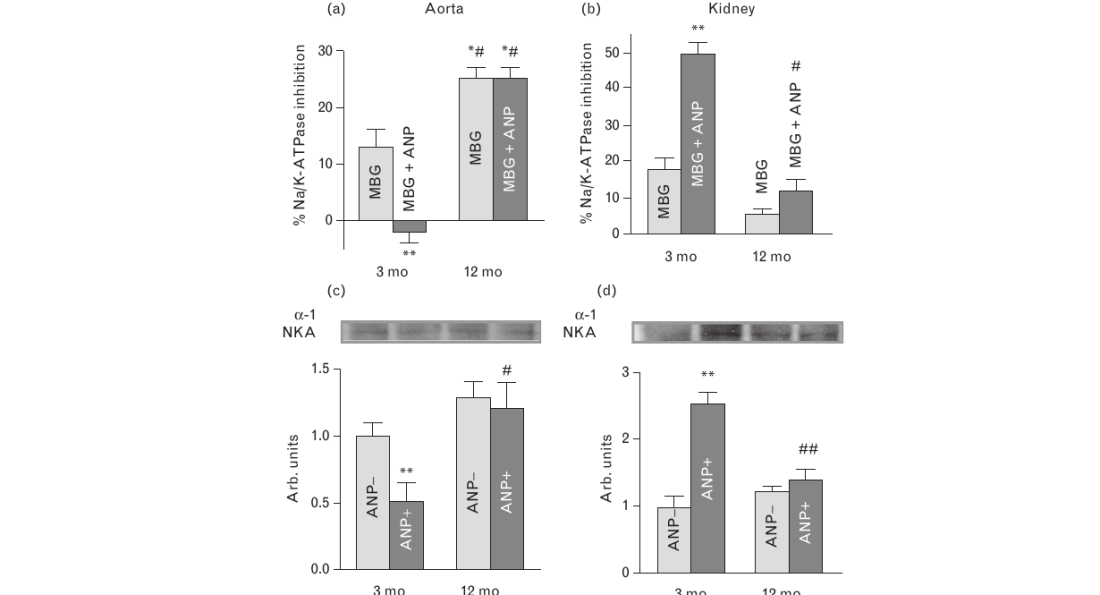
As demonstrated in Fig. 2a and b, baseline plasma and urinary levels of MBG were comparable in both older and young rats, but the response of plasma and urinary MBG to NaCl loading was greater in 12-month-old than in 3-month old rats. Baseline plasma levels of ANP and renal excretion of cGMP (Fig. 2c and d) were similar in both older and young animals. As presented in Fig. 2c and d, animals of both ages exhibited a comparable response of ANP and cGMP to NaCl loading. Data on the levels of expression of NKA isoforms, PKG isoforms, and PDE5 in aortic sarcolemma and in renal medulla are summarized in Figs 3 and 4. As compared with younger animals, the levels of α-1-NKA increased two-fold in aortic sarcolemma (Fig. 3a) and by 40% in the kidney (Fig. 3b) of 12-month-old rats. In aortic sarcolemma of the older animals, the levels of α-2-NKA were significantly reduced (Fig. 3d), whereas the levels of α-3-NKA were similar in both older and young rats (Fig. 3c). The levels ofPKG1 and PKG2 isoforms in vascular sarcolemma and renal medulla, respectively, were markedly reduced in older vs. young rats (Fig. 4a and b), whereas the levels of PDE5 were elevated in tissues of both older and younger rats (Fig. 4c and d).
In accord with our previous results [10,20], in-vitro MBG exhibited a concentration-dependent inhibition of the NKA in both aortic sarcolemma and renal medulla (Fig. 5). In both tissues, MBG-induced NKA inhibition occurred at the level of higher-affinity and lower-affinity binding (Table 2). As demonstrated in Fig. 5a, NKA from aortic sarcolemma of older animals exhibited heightened sensitivity to MBG as compared with that of younger animals. In the kidney, on the contrary, the sensitivity of NKA to MBG was reduced in 12-month-old vs. 3-month-old animals (Fig. 5b). Notably, in both tissues, age-dependent changes in MBG sensitivity of the NKA occurred at the level of higher-affinity, but not lower-affinity, binding (Table 2).
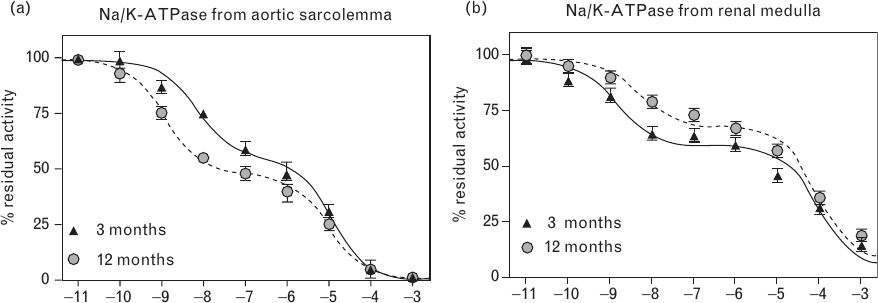
Table 2
Interaction of MBG with the Na/K-ATPase from aorta and renal medulla of Sprague-Dawley rats
| High-affinity IC50 (nM) | Low-affinity IC50 (μM) | |
|---|---|---|
| Aortic sarcolemma, 3-month-old | 6.9 ± 1.8 | 12 ± 2 |
| Aortic sarcolemma, 12-month-old | 1.2 ± 2.1 | 10 ± 2 |
| Renal medulla, 3-month-old | 1.4 ± 2.7 | 90 ± 3 |
| Renal medulla, 12-month-old | 5.0 ± 1.8 | 93 ± 3 |
Next (Fig. 6), we studied the modulatory effect of low concentration of α-hANP (1 nmol/l) on NKA inhibition induced by MBG at a concentration of 1 nmol/l, which is the concentration in plasma of rats with NaCl-induced hypertension [13,19], and on NKA from aortic sarcolemma and renal medulla in rats of both ages. Similar to that observed in our previous work [10], 1 nmol/l α-ANP alone did not significantly affect NKA activity in both aortic sarcolemma and renal medulla in rats of both ages (data not shown). As shown in Fig. 6a and b, in 3-month-old rats, 1 nmol/l α-ANP markedly reduced MBG-induced NKA inhibition in aortic sarcolemma but potentiated the effect of 1 nmol/l MBG on the NKA from outer medulla. In both tissues from aged rats, however, α-ANP failed to modulate NKA-inhibitory effect of low concentration of MBG. Thus, in older rats, in vitro, 1 nmol/l α ANP did not reduce the effect of MBG on NKA from aortic sarcolemma and did not potentiate MBG-induced NKA inhibition in renal medulla, as it did in younger rats. As demonstrated in Fig. 6c, in younger rats, 1 nmol/l α-ANP enhanced the levels of phosphorylation of α-1-NKA in renal medulla, but reduced its level in aortic sarcolemma. In aged rats, 1 nmol/l α-ANP did not affect the level of NKA phosphorylation in aortic sarcolemma and renal medulla (Fig. 6d).
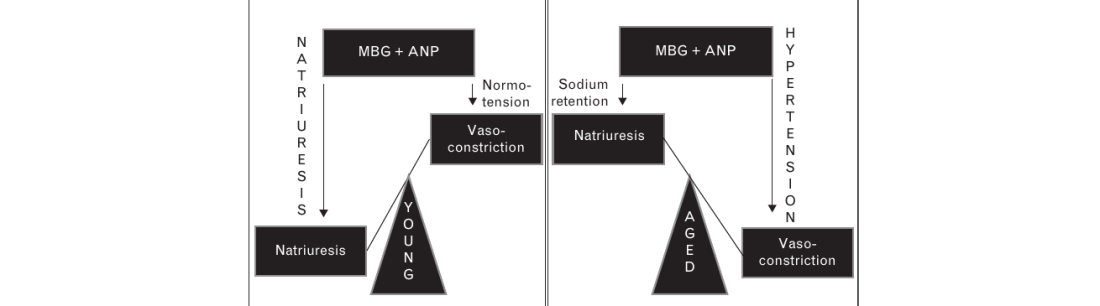
DISCUSSION
The main observation of the present study is that in normotensive rats, the development of age-associated salt sensitivity is accompanied by exaggerated response of MBG to NaCl loading occurring in the presence of markedly reduced levels of PKG in the vasculature and kidney. Downregulation of PKG in vascular smooth muscle and kidney and resultant reduction in cGMP-dependent signaling lead to the loss of the ability of ANP to modulate interactions between MBG and NKA. As a result, in the older animals ANP fails to potentiate MBG-induced inhibition of NKA in the kidney, leading to sodium retention, and fails to reduce MBG-induced inhibition of the NKA in the vasculature, leading to vasoconstriction. As schematically presented in Fig. 7, such a shift in ANP/PKG-dependent modulation of the effects of MBG would be expected to contribute to the development of salt sensitivity of blood pressure. Accordingly, following acute NaCl loading, 12-month-old rats exhibit a greater pressor response and a lesser natriuretic response to NaCl loading compared with 3-month-old rats. In the present study, the activity of the NKA in renal outer medulla in older rats was reduced, whereas the levels of α-1-NKA protein were higher in younger animals.
In the present study, the amount of sodium that was administered to the rats was above the physiological range, but it is unlikely that the changes in the levels of CTS occurred due to the changes in plasma osmolality. Previously, it has been demonstrated that in rats, concentrations of plasma CTS rose following a similar degree of NaCl loading, but did not change after hypertonic loading with sucrose [21]. Later, in Dahl-S rats, we demonstrated that the pattern of responses of CTS to acute NaCl challenge [20,22] was similar to that in Dahl-S rats chronically administered high NaCl [13] and in normotensive humans on a high NaCl intake [23].
Our present observations of reduced vascular and renal levels of PKG and impaired PKG-dependent modulation of NKA activity observed in older rats agree with the previous finding that in aging rats, a decrease in cGMP/PKG modulation underlies reduction of NKA levels in the trachea and brain [24]. In accordance with previous reports, in the present study, we detected the presence of α-1-NKA, α-2-NKA, and α-3-NKA isoforms in aorta, whereas α-1-NKA was present exclusively in renal medulla [25]. Our present observation of age-associated upregulation of α-1-NKA in renal medulla and reduction of NKA activity in aorta confirms previous findings of increased α-1-NKA protein in the medulla of aged rats [26] and age-related decline in the activity of the sodium pump in the aorta and tail artery [27]. Notably, in the present study, in older rats that exhibited salt sensitivity of blood pressure, the levels of MBG-sensitive α-1-NKA were elevated in both renal medulla and aortic sarcolemma, whereas NKA activity in both tissues was reduced in younger animals. This pattern is similar to that previously observed in left ventricular myocardium in Dahl-S rats, in which the development of salt-induced hypertension was associated with increased cardiac levels of α-1-NKA protein and reduced NKA activity in left ventricular sarcolemma [28]. In that study, the reduction of NKA activity has been attributed to protein kinase C-induced sensitization of the NKA to MBG [28]. Whether similar mechanism is implicated in pathogenesis of age-associated hypertension and whether our observations made in aorta are relevant to the resistant arteries remains to be studied.
Recently, we studied the effect of acute NaCl loading on the responses of MBG, ANP, and cGMP, and on the pattern of inhibition of renal and vascular sodium pump in young Sprague–Dawley rats and Dahl-S rats [14]. In that study, Dahl-S rats, in which salt sensitivity was determined genetically, exhibited a pattern of sodium pump inhibition similar to that observed in the older Sprague–Dawley rats in the present study, that is, greater inhibition of the sodium pump in the vasculature in the presence of reduced sodium pump inhibition in the kidney following acute NaCl loading [14]. Thus, rats with genetically determined salt sensitivity and those with age-associated salt sensitivity exhibited impaired cGMP/PKG-dependent signaling and a similar pattern of responses of renal and vascular sodium pump to NaCl loading. In Dahl-S rats, however, the level of this impairment occurred upstream from that in older Sprague–Dawley rats. Although in the former both ANP and cGMP responses to NaCl loading were blunted, in the latter, responses of ANP and cGMP to NaCl challenge were unaltered, but marked reduction in the levels of PKG along with an increase in the levels of PDE5 impaired cGMP/PKG dependent signaling.
Aperia et al. [9] hypothesized that several hormones (dopamine, ANP, angiotensin, and norepinephrine) implicated in regulation of natriuresis, and acting through a signaling cascade involving activation of second messengers and protein kinases, are capable of modulating renotubular NKA activity by affecting its state of phosphorylation. Our previous [10] and present data, demonstrating that ANP enhanced the levels of α-1-NKA phosphorylation in the kidney and exhibited an opposite effect in the vasculature, agree with this hypothesis. Although addressed in several previous studies, the molecular mechanisms underlying the effect of PKG on NKA phosphorylation are not well understood. Scavone et al. [16] demonstrated that low ANP concentrations, acting via cGMP-dependent mechanism, inhibit the NKA from rat renal medulla. As this cGMP-dependent effect has been mimicked by a protein phosphatase inhibitor, okadaic acid, these authors concluded that sodium pump phosphorylation might be a target for cGMP-dependent effects of ANP [16]. Fotis et al. [29] demonstrated that PKG phosphorylates α-isoform of the sodium pump from various species, and that the site for PKG phosphorylation is located in the central part of the a-subunit polypeptide chain, unlike phosphorylation sites for protein kinases A and C located at the NKA a-subunit C and A terminus, respectively. Another possibility is that the effects of PKG on the sodium pump could be mediated by phosphorylation of DARPP-32, a substrate for PKG and protein kinase A, which exerts its effect on renal NKA phosphorylation and activity via dephosphorylation of protein phosphatase-1 [30].
In conclusion, in Sprague–Dawley rats, the development of age-associated salt sensitivity of blood pressure is accompanied by a reduced responsiveness of renal sodium pump to MBG, whereas vascular sodium pump exhibits heightened MBG sensitivity. Downregulation of PKG1 and PKG2 in the vasculature and in the kidney, respectively, and a resultant loss of the ability of ANP to potentiate MBG-induced inhibition of renal NKA and to offset the effect of MBG on vascular sodium pump appear to underlie these effects. These data agree with a recent finding that a cluster of single nucleotide polymorphisms located in the PKG1 gene associates with the salt sensitivity of blood pressure in hypertensive humans [31]. Our findings indicate that MBG, acting via NKA-dependent signaling, stimulates the synthesis of collagen in cardiac, renal, and vascular tissues [32], and that immunoneutralization of MBG reverses cardiac fibrosis in rats with renal failure [33]. Because fibrosis is a hallmark of cardiovascular aging [34], and because natriuretic peptides exhibit antifibrotic effects [35], the impact of age-associated reduction in cGMP/PKG signaling on the profibrotic effects of CTS merits further studies.
References
| # | Year | Authors | Title |
|---|---|---|---|
| 1 | 1990 | Khaw KT, Barrett-Connor E | Increasing sensitivity of blood pressure to dietary sodium and potassium with increasing age. A population study using casual urine specimens. Am J Hypertens.. 3, 505-511. |
| 2 | 2001 | Sacks FM, Svetkey LP, Vollmer MW, Appel LJ, Bray GA, Harsha D | Effects on blood pressure of reduced dietary sodium and the dietary approaches to stop hypertension (DASH) diet. N Engl J Med.. 344, 3-10. |
| 3 | 1997 | Cowley AW | Genetic and nongenetic determinants of salt sensitivity and blood pressure. Am J Clin Nutr.. 65, 587S-593S. |
| 4 | 2009 | Bagrov AY, Shapiro JI, Fedorova OV | Endogenous cardiotonic steroids: physiology, pharmacology and novel therapeutic targets. Pharmacol Rev.. 61, 9-38. |
| 5 | 2002 | Bełtowski J, Wócicka G | Regulation of renal tubular sodium transport by cardiac natriuretic peptides: two decades of research. Med Sci Monit.. 8, RA39-RA52. |
| 6 | 2007 | Richards AM | Natriuretic peptides: update on Peptide release, bioactivity, and clinical use. Hypertension.. 50, 25-30. |
| 7 | 2000 | Bailly C | Effect of luminal atrial natriuretic peptide on chloride reabsorption in mouse cortical thick ascending limb: inhibition by endothelin. J Am Soc Nephrol.. 11, 1791-1797. |
| 8 | 1994 | Florkowski CM, Richards AM, Espiner EA, Yandle TG, Frampton C | Renal, endocrine, and hemodynamic interactions of atrial and brain natriuretic peptides in normal men. Am J Physiol.. 266, R1244-R1250. |
| 9 | 1994 | Aperia A, Holtback U, Syren ML, Svensson LB, Fryckstedt J, Greengard P | Activation/deactivation of renal Na/K-ATPase: a final common pathway for regulation of natriuresis. FASEB J.. 8, 436-469. |
| 10 | 2006 | Fedorova OB, Agalakova NI, Morrell CH, Lakatta EG, Bagrov AY | ANP differentially modulates marinobufagenin-induced sodium pump inhibition in kidney and aorta. Hypertension.. 48, 1160-1168. |
| 11 | 2003 | Hebert SC | Bartter syndrome. Curr Opin Nephrol Hypertens.. 12, 527-532. |
| 12 | 2002 | Fridman AI, Matveev SA, Agalakova NI, Fedorova OV, Lakatta EG, Bagrov AY | Marinobufagenin, an endogenous ligand of a-1 Na/K-ATPase, is a marker of congestive heart failure severity. J Hypertens.. 20, 1189-1194. |
| 13 | 2002 | Fedorova OV, Talan MI, Agalakova NI, Lakatta EG, Bagrov AY | An endogenous ligand of a-1 sodium pump, marinobufagenin, is a novel mediator of sodium chloride dependent hypertension. Circulation.. 105, 1122-1127. |
| 14 | 2009 | Bagrov AY, Agalakova NI, Kashkin VA, Fedorova OV | Endogenous cardiotonic steroids and differential patterns of sodium pump inhibition in NaCl-loaded salt-sensitive and normotensive rats. Am J Hypertens.. 22, 559-563. |
| 15 | 2001 | Lin CS, Liu X, Tu R, Chow S, Lue TF | Age-related decrease of protein kinase G activation in vascular smooth muscle cells. Biochem Biophys Res Commun.. 287, 244-248. |
| 16 | 2005 | Scavone C, Munhoz CD, Kawamoto EM, Glezer I, de Sá Lima L, Marcourakis T, Markus RP | Age-related changes in cyclic GMP and PKG-stimulated cerebellar Na,K-ATPase activity. Neurobiol Aging.. 26, 907-916. |
| 17 | 2009 | Cui R, Iso H, Yamagishi K, Ohira T, Tanigawa T, Kitamura A | Relationship of urinary cGMP excretion with aging and menopausal status in a general population. J Atheroscler Thromb.. 16, 457-462. |
| 18 | 1997 | Feschenko MS, Sweadner KJ | Phosphorylation of Na,K-ATPase by protein kinase C at Ser18 occurs in intact cells but does not result in direct inhibition of ATP hydrolysis. J Biol Chem.. 272, 17726-17733. |
| 19 | 2008 | Fedorova OV, Simbirtsev AS, Kolodkin NI, Kotov AY, Agalakova NI, Kashkin VA | Monoclonal antibody to an endogenous bufadienolide, marinobufagenin, reverses preeclampsia-induced Na/KATPase inhibition in lowers blood pressure in NaCl-sensitive hypertension. J Hypertens.. 26, 2414-2425. |
| 20 | 2000 | Fedorova OV, Lakatta EG, Bagrov AY | Differential effects of acute NaCl loading on endogenous ouabain-like and marinobufagenin-like ligands of the sodium pump in Dahl hypertensive rats. Circulation.. 102, 3009-3014. |
| 21 | 1997 | Yamada K, Goto A, Nagoshi H, Terano Y, Omata M | Elevation of ouabain-like compound levels with hypertonic sodium chloride load in rat plasma and tissues. Hypertension.. 30, 94-98. |
| 22 | 2005 | Fedorova OV, Agalakova NI, Talan MI, Lakatta EG, Bagrov AY | Brain ouabain stimulates peripheral marinobufagenin via angiotensin II signalling in NaCl loaded Dahl-S rats. J Hypertens.. 23, 1515-1523. |
| 23 | 2008 | Anderson DE, Fedorova OV, Morrell CH, Kashkin VA, Bagrov AY, Lakatta EG | Endogenous sodium pump inhibitors are linked to age associated increases in salt sensitivity of blood pressure in normotensives. Am J Physiol Regul Integr Physiol.. 294, R1248-R1254. |
| 24 | 2008 | Kawamoto EM, Munhoz CD, Lepsch LB, de Sá Lima L, Glezer I, Markus RP | Age-related changes in cerebellar phosphatase-1 reduce Na,K-ATPase activity. Neurobiol Aging.. 29, 1712-1720. |
| 25 | 1998 | Blanco G, Mercer RW | Isozymes of the Na-K-ATPase: heterogeneity in structure, diversity in function. Am J Physiol.. 275, F633-F650. |
| 26 | 2010 | Silva E, Pinto V, Simao S, Serrao MP, Afonso J, Amaral J | Renal aging in WKY rats: changes in Na+,K+-ATPase function and oxidative stress. Exp Gerontol.. 45, 977-983. |
| 27 | 1988 | Hennessy JF | Vascular tissue (Na,K)ATPase activity and aging in the F344 rat. Mech Ageing Dev.. 43, 153-159. |
| 28 | 2003 | Fedorova OV, Talan MI, Agalakova NI, Droy-Lefaix M-T, Lakatta EG, Bagrov AY | Reduction in myocardial PKC b2, Na/K-ATPase sensitivity to marinobufagenin and blood pressure in response to cicletanine. Hypertension.. 41, 505-511. |
| 29 | 1999 | Fotis H, Tatjanenko LV, Vasilets LA | Phosphorylation of the alpha subunits of the Na,ATPase from mammalian kidneys and Xenopus oocytes by cGMP-dependent protein kinase results instimulation of ATPase activity. Eur J Biochem.. 260, 904-910. |
| 30 | 2001 | Eklof AC, Holtback U, Svennilson J, Fienberg A, Greengard P, Aperia A | Increased blood pressure and loss of ANP-induced natriuresis in mice lacking DARPP-32 gene. Clin Exp Hypertens.. 23, 449-460. |
| 31 | 2011 | Citterio L, Simonini M, Zagato L, Salvi E, Delli Carpini S, Lanzani C | Genes involved in vasoconstriction and vasodilation system affect saltsensitive hypertension. PLoS One.. 6, e19620. |
| 32 | 2010 | Fedorova OV, Shapiro JI, Bagrov AY | Endogenous cardiotonic steroids and salt-sensitive hypertension. Biochim Biophys Acta.. 1802, 1230-1236. |
| 33 | 2012 | Haller ST, Kennedy DJ, Shidyak A, Budny GV, Malhotra D, Fedorova OV | Monoclonal antibody against marinobufagenin reverses cardiac fibrosis in rats with chronic renal failure. Am J Hypertens.. 25, 690-696. |
| 34 | 2008 | Susic D, Frohlich ED | The aging hypertensive heart: a brief update. Nat Clin Pract Cardiovasc Med.. 5, 104-110. |
| 35 | 2006 | Nishikimi T, Maeda N, Matsuoka H | The role of natriuretic peptides in cardioprotection. Cardiovasc Res.. 69, 318-328. |