Abstract
Endogenous cardiotonic steroids (CTS), also called digitalis-like factors, have been postulated to play important roles in health and disease for nearly half a century. Recent discoveries, which include the specific identification of endogenous cardenolide (endogenous ouabain) and bufadienolide (marinobufagenin) CTS in humans along with the delineation of an alternative mechanism by which CTS can signal through the Na+/K+-ATPase, have increased the interest in this field substantially. Although CTS were first considered important in the regulation of renal sodium transport and arterial pressure, more recent work implicates these hormones in the regulation of cell growth, differentiation, apoptosis, and fibrosis, the modulation of immunity and of carbohydrate metabolism, and the control of various central nervous functions and even behavior. This review focuses on the physiological interactions between CTS and other regulatory systems that may be important in the pathophysiology of essential hypertension, preeclampsia, endstage renal disease, congestive heart failure, and diabetes mellitus. Based on our increasing understanding of the regulation of CTS as well as the molecular mechanisms of these hormone increases, we also discuss potential therapeutic strategies.
I. Introduction
The topic of this review is the digitalis-like factors, which are also referred to as inhibitors of the Na+/K+ATPase (de Wardener and Clarkson, 1985; Goto et al., 1992; Schoner, 1992) or endogenous cardiotonic steroids (CTS1). As we will discuss, these CTS link dietary NaCl and cardiovascular and renal disease. Although the importance (and the very existence) of such factors has been a matter of controversy (Kelly and Smith, 1992; Hansen, 2003), remarkable progress has been achieved during the past 15 years. These breakthroughs are illustrated in a series of articles and include 1) positive identification of specific CTS in experimental animals and humans (Hamlyn et al., 1991; Lichtstein et al., 1993; Bagrov et al., 1998; Komiyama et al., 2005), 2) establishment of alterations in concentrations as well as the role(s) of CTS in animal models and human disease states (Ferrandi et al., 2005; Haddy, 2006; Huang et al., 2006; Schoner and Scheiner-Bobis, 2007), and, in parallel, 3) the discovery of cell signaling functions of the Na+/K+-ATPase and its involvement in many aspects of basic cell biology (Xie and Askari, 2002; Wasserstrom and Aistrup, 2005; Orlov and Hamet, 2006; Nesher et al., 2007; Schoner and Scheiner-Bobis, 2007). The main goals of the present review are to emphasize the clinical implications of CTS in human health and disease and to demonstrate potential targets for new therapies.
Abbreviations: CTS, Cardiotonic steroid(s); TM, transmembrane; MBG, marinobufagenin; PKC, protein kinase C; ANP, atrial natriuretic peptide; GFR, glomerular filtration rate; EGFR; epidermal growth factor receptor; EO, endogenous ouabain; ACTH, adrenocorticotrophin; Dahl-S, Dahl salt-sensitive rats; PST2238, rostafuroxin; BP, blood pressure; RAS, renin-angiotensin system; CHF, congestive heart failure; ATII, Angiotensin II; AT, angiotensin; ERK, extracellular signal-regulated kinase; PI(3)K, phosphoinositide 3-kinase; ROS, reactive oxygen species; NHE3, Na+/H+ exchanger isoform 3; ESRD, end-stage renal disease; LV, left ventricular; α-hANP, α-human atrial natriuretic peptide; OSA, obstructive sleep apnea; PaCO2, arterial tension of carbon dioxide; PKG, protein kinase G; NCX1, Na+/Ca2+ exchanger 1; SEA0400, [2-[4-[(2,5-difluorophenyl)methoxy]phenoxy]-5-ethoxyaniline]; KB-R7942, 2-[2-[4-(4-nitrobenzyloxy)phenyl]ethyl]isothiourea methanesulfonate.
II. Na+/K+-ATPase
A. Structure and Function of the Na+/K+-ATPase
The discovery of the sodium pump was a critical step in the 300-year study of the cell as a basic unit of animal life. More importantly, the sodium pump gave substance to the concept of the cell membrane, which isolates the milieu interior from the external environment and/or the environment of other cells. Based on the asymmetrical distribution of sodium and potassium ions, the scientific community was ready to accept the existence of “submicroscopic pumps, installed across the cell membrane,” which could actively participate in “fine-tuning” of the transmembrane ion gradients in accordance with changes in the physiological needs of cells (Ling, 2007).
The discovery of the sodium pump is generally credited to Skou (1957) for his experiments with crab nerve homogenate that clearly demonstrated the existence of a protein-based structure, incorporated in the cell membrane, which pumped sodium ions outside and potassium ions inside living cells, and in so doing, converted chemical energy into work. It is noteworthy that this discovery was possible because of the existence of ouabain, a specific sodium pump inhibitor of steroidal nature and plant origin, which later was found to be identical to one of the endogenous mammalian inhibitors of activity of sodium pump (Hamlyn et al., 1982, 1991).
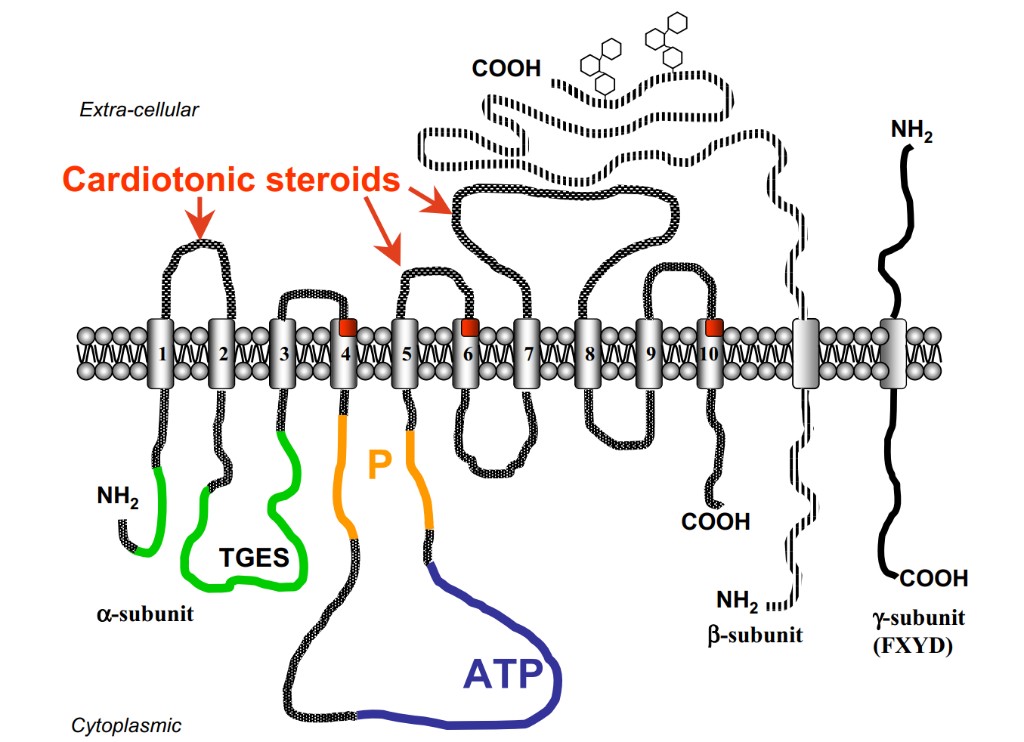
The sodium pump, or Na+/K+-ATPase [(Na+ + K+)stimulated adenosine triphosphatase; EC 3.6.3.9], is an active transport system of sodium and potassium ions that is highly conserved in all eucaryote cells. It is a α subunit has 10 transmembrane segments. It contains the binding sites for Na+ and CTS on the extracellular segments and the binding sites for K+ and ATP on the intracellular loops. Asp369, in particular, is critical for the binding of a phosphate group and is an essential site for ATP hydrolysis (Lingrel, 1992). Two more residues, Lys501 and Asp586, may also participate in the binding of ATP (Farley et al., 1997) (Fig. 1). The regulatory β subunit is a single-transmembrane protein with a glycosylation site controlling the activity of Na+/K+-ATPase (Blanco and Mercer, 1998). At present, the αβ complex is believed to form the functional dimeric units of the sodium pump. These dimers may also be organized in tetramers or possibly stack into ribbons (Söderholm et al., 1988).
According to the Albers-Post model, the Na+/K+-ATPase “pumps” sodium ions from the inside of the cell to the outside while moving potassium ions in the opposite direction, both against existing concentration gradients in an energy-dependent process. The α subunit, in the presence of Na+ and Mg2+, is phosphorylated by ATP, followed by occlusion of three cytosolic Na+ ions. This high-energy E1P form of the enzyme, loaded with Na+ P
B. Na+/K+-ATPase as a Specific Receptor for Cardiotonic Steroids
whereas ouabain is 3 orders of concentration less active with renal Na+/K+-ATPase (Fedorova et al., 2000, 2001b).
The sensitivity of the sodium pump to cardiotonic steroids is controlled by multiple mechanisms in addition to the tissue specificity of α and β isoforms distributions. Thus, it is known that protein kinases phosphorylate the Na+/K+-ATPase in a tissueand isoformspecific fashion (Bertorello and Katz, 1993; Blanco and Mercer, 1998; Therien and Blostein, 2000). The α subunit is phosphorylated by protein kinase A and protein kinase C (PKC) (Bertorello et al., 1991); PKC directly phosphorylates Ser18 on the intracellular domain (Feschenko and Sweadner, 1997; Feschenko et al., 1997) of the α1 isoform of Na+/K+-ATPase in vascular smooth muscle from human mesenteric artery, which in turn causes increased sensitivity of sodium pump to MBG but not to ouabain (Fedorova et al., 2002a). Some studies indicate that dopamine-induced phosphorylation of the α subunit at Ser18 results in a clathrin-dependent internalization of the sodium pump (Chibalin et al., 1999).
Another mechanism of modulation of sodium pump activity involves the cGMP-sensitive phosphorylation of Na+/K+-ATPase; this may be initiated by atrial natriuretic peptide (ANP). It is noteworthy that whereas cGMP induces Na+/K+-ATPase phosphorylation in renal epithelial cells, it actually reduces levels of Na+/K+ATPase phosphorylation in smooth muscle cells (Fedorova et al., 2006).
The three classic features of Na+/K+-ATPase (the pump, the enzyme, and the receptor to cardiotonic steroids) are being understood in substantially greater detail, and, in some aspects, have undergone a paradigm shift in understanding. In particular, there is evidence that extremely low concentrations of cardiotonic steroids, which are unlikely to inhibit the enzymatic function of the sodium pump (Akimova et al., 2005, 2008a; Orlov and Hamet, 2006), are able to initiate several signaling pathways, which may be extremely important for a variety of cell functions. This process is discussed in detail in section V. Szent-Gyorgyi (1953) was one of the first to hypothesize that digitalis drugs, are, in fact, a substitute of endogenous compounds participating in the regulation of cardiac muscle contractility. Thus, the further discovery of Na+/K+-ATPase with a receptor site for ouabain became a landmark in the discovery of the endogenous CTS.
III. Endogenous Digitalis
A. Early Evidence
For many years, increased dietary NaCl intake has been suspected to increase the risk of cardiovascular and renal diseases (Ritz, 1996; Meneton et al., 2005). However, during the several last decades, the importance of dietary NaCl for development of hypertension has shifted from a topic of debate to a well established phenomenon (De Wardener and MacGregor, 2002; Ritz et Although digitalis-like substances were almost certainly a part of herbal remedies administered as long ago as the Roman empire, most authorities agree that serious scientific study and medicinal application began with William Withering’s observation of a salutary effect of foxglove (Digitalis purpurea). Withering, a British physician and herbalist, apparently obtained foxglove initially from “an old woman in Shropshire,” who had apparently administered a variety of herbal remedies to local patrons.
In his 1785 monograph, “An Account of the Foxglove and Some of Its Medical Uses; With Practical Remarks on Dropsy and Other Diseases,” Withering detailed nearly a decade of studying both the beneficial and toxic effects of foxglove. In 1869, Claude Adolphe Nativelle, a French pharmacist isolated a material he called “digitalin” from the foxglove plant. Six years later, the German chemist, Oswald Schmiedeberg, isolated the first pure glycoside in crystal form from foxglove, which he called “digitoxin” (Lesney, 2002). CTS belonging to the class of bufadienolides were also introduced into routine clinical practice more than 1000 years ago. The dried skin of toads, which contains various bufadienolide derivatives, comprises one of the main active principles of traditional Chinese medicine Sen-So (or Ch’an Su), and has been used in the treatment of cardiac dysfunction (Chen and Kovaríková, 1967). The pharmacological effects of digitalis and related compounds were reviewed in many excellent articles (Braunwald and Klocke, 1965; Langer, 1972; Repke, 1972; Smith and Haber, 1973; Lindenmayer, 1976; Akera and Brody, 1977; Gillis and Quest, 1979; McDonough et al., 1995).
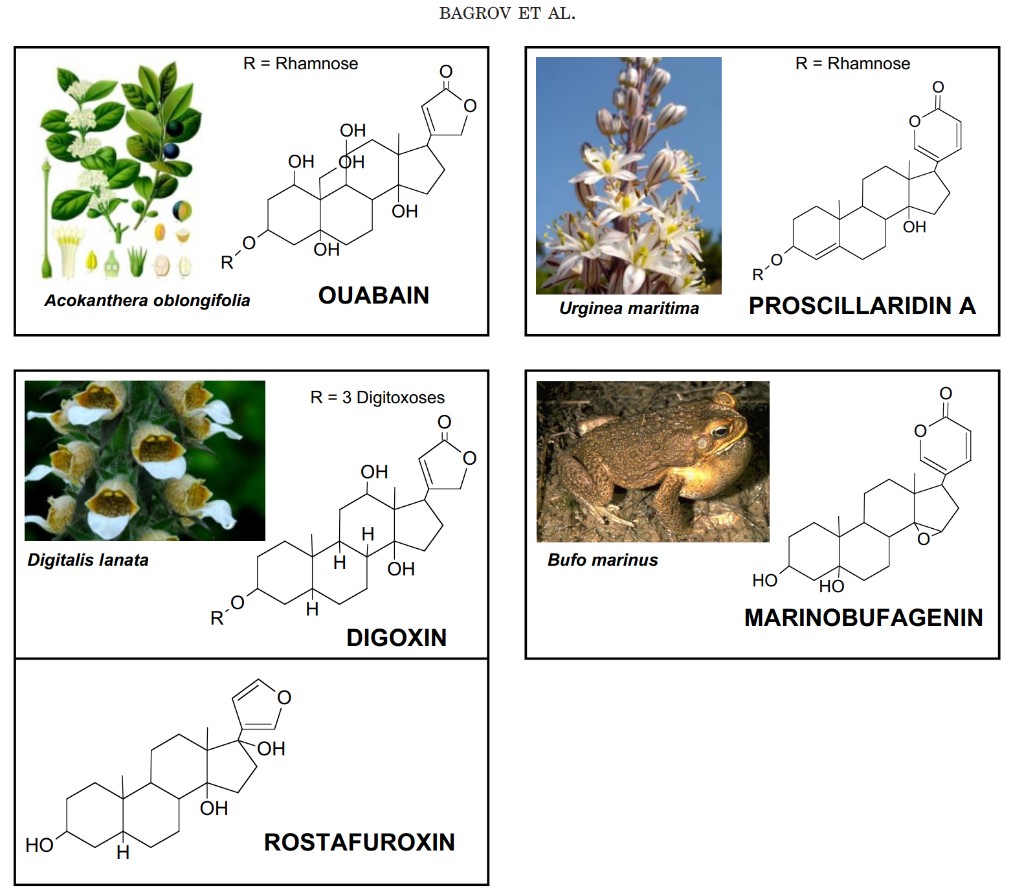
Digitalis-like inhibitors, or CTS, have a specific binding site on the extracellular loops (TM1–TM2, TM5–TM6, and TM7–TM8) of the α subunit of the Na+/K+-ATPase. Some amino acids from the transmembrane regions (M4, M6, and M10) also interact with ouabain; this suggests that the hydrophobic regions of CTS may actually be inserted in the membrane inside which they interact with the α subunit of sodium pump (Burns et al., 1996; Croyle et al., 1997; Vasilets et al., 1998). Amino acids from 111 to 122 sequences (extracellular TM1–TM2 loop) form the most important part of the putative CTS binding site (Fig. 1). It is noteworthy that the dissimilarities in the amino acid sequence between different species and different isoforms underlie the different sensitivity of the sodium pump to a variety of cardiotonic steroids (Blanco et al., 1999; Mobasheri et al., 2000; Geering, 2005, 2006), which include cardenolides and bufadienolides (this issue is reviewed in detail in sections IV.A and IV.B) (Fig. 2). For example, rodent renal epithelial Na+/K+-ATPase, consisting nearly exclusively of the α1 isoform, may be inhibited by a bufadienolide marinobufagenin (MBG) at nanomolar concentration range, R = Rhamnose R = Rhamnose R R Acokanthera oblongifolia Urginea maritima PROSCILLARIDIN A R = 3 Digitoxoses Digitalis lanata R
B. Regulation of Volume and the Third Factor
The Third Factor was a topic of great interest in the 1960s and 1970s (Guyton et al., 1980). Cort and Lichardus (1963) demonstrated that a circulating substance in cats subjected to carotid artery occlusion induced natriuresis in rats and inhibited sodium transport in frog skin. Buckalew et al. (1970) demonstrated that an ultrafiltrate of volume-expanded dogs inhibited sodium transport in toad bladders and postulated that this ultrafiltrate contained an inhibitor of the Na+/K+ATPase. Extremely important contributions were made
C. Hypertension, Dietary NaCl Intake, and Concept of Natriuretic Hormone
Natriuretic Hormone Despite the excitement of the 1960s and 1970s, enthusiasm for the concept of Na+/K+-ATPase inhibitors as third factors decreased during the 1980s and 1990s in part because of inconsistencies with the experimental data. Most assays were based on cross-reactivity to the medication digoxin, and such cross-reactivity to other CTS varied dramatically (Sheiner et al., 1974; Bergdahl et al., 1979; Gruber et al., 1980; Bergdahl and Molin, 1981; Kelly, 1986; Pleasants et al., 1986). However, the most important inconsistency was that digitalis, the prototypical inhibitor of the Na+/K+-ATPase was not natriuretic in normal subjects (Hauptman and Kelly, 1999).
In addition, atrial (and brain) natriuretic peptide(s) were discovered; these hormones clearly were natriuretic and could be easily measured, and their circulating concentrations were obviously increased by volume expansion (de Bold et al., 1981; Agnoletti et al., 1987; Bruneau et al., 1997; Lee and Burnett, 2007).
Interest in the study of CTS has steadily increased in the past decade for several reasons. First and perhaps foremost, multiple endogenous digitalis-like factors have been isolated and chemically characterized in experimental animals and humans (Hamlyn et al., 1991; Lichtstein et al., 1993; Bagrov et al., 1998; Komiyama et al., 2005). Specifically, ouabain, MBG, telocinobufagin, and bufalin have each been unequivocally identified in human plasma and/or urine. Also quite importantly, a signal cascade that does not seem to depend on enzymatic inhibition of the Na+/K+ATPase but rather occurs when CTS bind caveolar Na+/K+-ATPase in the company of Src and EGFR has been identified (Xie and Askari, 2002; Liu et al., 2003; Wang et al., 2004; Pierre and Xie, 2006; Liang et al., 2007). The measurement of CTS has evolved from bioassays and cross-reaction with digitalis because of the definite identification of CTS and the development of high-quality immunoassays. At present, there is a commercially available assay for ouabain, whereas measurements of MBG (Fedorova et al., 2002b, 2008), telocinobufagin, and bufalin still require research assays. Plasma levels of ouabain and MBG in humans seem to be in the range of 200 to 1500 pM in health and disease.
IV. Salt-Sensitive Hypertension, Concept of Natriuretic Hormone, and Elucidation of Nature and Roles of Endogenous Cardiotonic Steroids
A. Endogenous Cardenolides
1. Endogenous Ouabain
reported that human plasma contains CTS indistinguishable from ouabain (Hamlyn et al., 1991; Ludens et al., 1991) (Fig. 2). Since that time, endogenous ouabain (EO) has also been isolated from bovine adrenal gland (Schneider et al., 1998), bovine hypothalamus (Tymiak et al., 1993), and rat adrenomedullary cells (Komiyama et al., 2001). Using NMR and mass spectrometry techniques, mammalian EO was found to be identical to the plant-derived ouabain (Hamlyn et al., 1991; Tymiak et al., 1993; Schneider et al., 1998). The adrenal cortex and hypothalamus are considered to be the sites of EO production (Komiyama et al., 2001; el-Masri et al., 2002; Murrell et al., 2005).
ACTH, angiotensin II, vasopressin, and phenylephrine were shown to facilitate the in vitro release of EO from adrenal cortex (Laredo et al., 1997; Shah et al., 1999).
Evidence indicates that EO does not fulfill the criteria for a putative natriuretic hormone but does play a role in the adaptation to both sodium depletion and sodium loading. Although some studies of salt loading in normotensive rats demonstrate increases in plasma [EO] (Butt et al., 1997; Ho et al., 1997), results of many other experiments performed in dogs (Ludens et al., 1993; Bagrov et al., 1996b), rats (Fedorova et al., 2001a,b), and humans (Manunta et al., 2001; Balzan et al., 2005) are contradictory. In hypertensive subjects, results are similarly inconsistent. In 180 untreated hypertensive pa by Schrier and de Wardener in the measurement and implications of this factor (De Wardener et al., 1961; de Wardener et al., 1968, 1971; Schrier et al., 1968a,b; Schrier, 1974). Kramer and Gonick (1974) were later able to demonstrate that volume expansion in rats produced a substance that could inhibit ATPase activity in rat kidney. Bricker and coworkers incorporated the concept of a circulating Na+/K+-ATPase inhibitor in a “trade-off” schema for renal failure progression and the pathogenesis of the uremic syndrome (Bourgoignie et al., 1970; Kaplan et al., 1974; Schmidt et al., 1974; Bricker et al., 1975). Although some of the effects attributed to a Third Factor are, in fact, caused by several factors in addition to aldosterone and GFR (e.g., atrial and brain natriuretic factors), this work initiated the search for these factors, of which cardiotonic steroids feature prominently.
Haddy and Overbeck (1976) and Overbeck et al. (1976) discovered that the Na+/K+ pump activity (ouabain-sensitive 86Rb uptake) in arteries and veins of dogs with low renin hypertension is decreased and that Na+/K+-ATPase activity in cardiac microsomes from rats with low renin hypertension is inhibited. Gruber et al. (1980) demonstrated that plasma volume expansion is associated with elevated levels of circulating digoxin-like immunoreactive material in dogs. Shortly thereafter, Hamlyn et al. (1982) demonstrated that plasma Na+/K+-ATPase inhibitory correlated positively with blood pressure in patients with essential hypertension. Kojima et al. (1982) showed that anti-digoxin antibody lowers blood pressure in rats with deoxycorticosterone-salt-induced hypertension. Taken together, we argue that these studies strongly implicated CTS in the pathogenesis of hypertension.
hormone. In rats, chronic peripheral administration of low doses of ouabain (10 –50 g/kg/day) has been reported to increase arterial pressure and to induce cardiac hypertrophy (Rossoni et al., 2002, 2006; Ferrandi et al., 2004; Briones et al., 2006; Cheung et al., 2006; Dostanic-Larson et al., 2006). Padilha et al.
(2008) have shown in rats that chronic administration of a low dose of ouabain is associated with an increase in the expression of cyclooxygenase-2 and with impairment of the release of endothelium-derived hyperpolarizing factor.
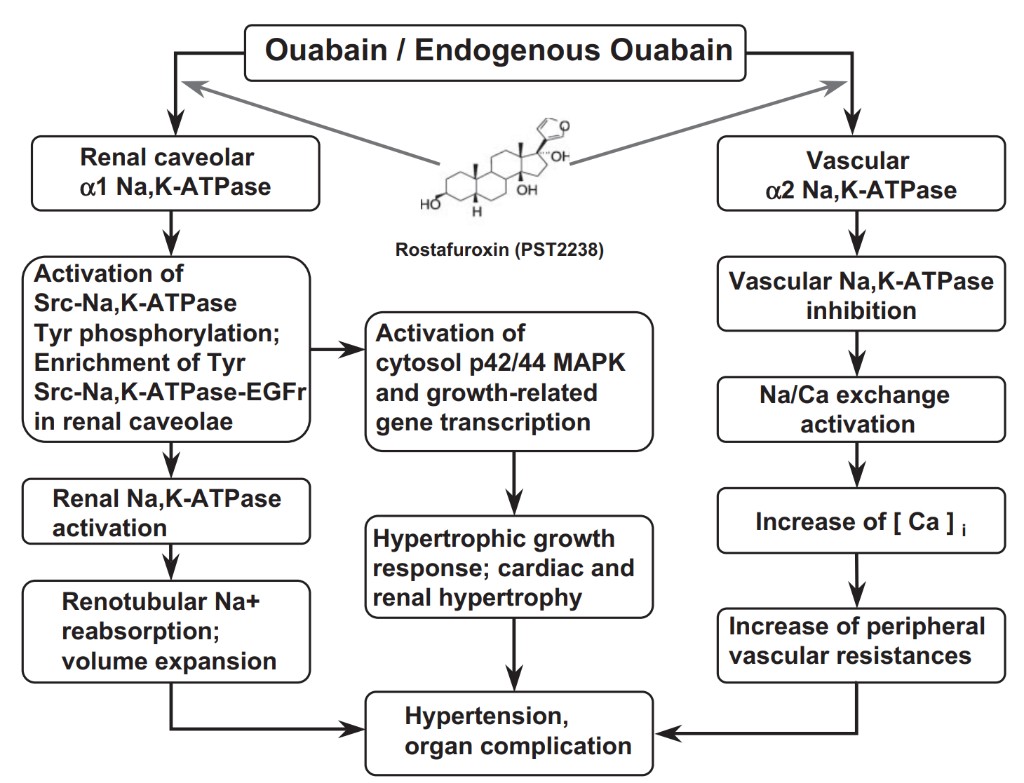
Three mechanisms, the “adducin paradigm,” the existence of highly sensitive ouabain-binding sites in vascular smooth muscle, and central effects, were proposed to link EO to vasoconstriction in hypertension (Fig. 3). A mechanism for the prohypertensive effect of EO has been suggested by experiments performed in Milan hypertensive rats, which carry a mutation of a gene of a cytoskeletal protein, adducin (Bianchi et al., 1994). In this strain of rats, both increased levels of EO and adducin mutation are associated with heightened expression and Na+/K+-ATPase activity in the renotubular epithelium. This heightened expression is apparently Ouabain / Endogenous Ouabain Renal caveolar α1 Na,K-ATPase Vascular α2 Na,K-ATPase Rostafuroxin (PST2238) Activation of Src-Na,K-ATPase Tyr phosphorylation; Enrichment of Tyr Src-Na,K-ATPase-EGFr in renal caveolae Renal Na,K-ATPase activation Renotubular Na+ reabsorption; volume expansion Activation of cytosol p42/44 MAPK and growth-related gene transcription Hypertrophic growth response; cardiac and Vascular Na,K-ATPase inhibition activation Increase of [ Ca ] i Increase of peripheral vascular resistances tients, Manunta et al. (2001) demonstrated that plasma [EO] did not change during volume expansion but rather increased after 2 weeks of sodium depletion. In a more recent study, in 13 healthy individuals, Manunta et al.
(2006) demonstrated that NaCl depletion produced a 4-fold elevation in plasma [EO], but in the same study, 3 days of NaCl loading (171 mEq of NaCl/day) caused a transient 13-fold elevation in plasma [EO]. A similar pattern of EO response (i.e., elevation followed by decrease) has been observed in Dahl salt-sensitive (Dahl-S) rats after acute and chronic NaCl loading (Fedorova et al., 2000, 2002b) and in normotensive human subjects during 6 days of high NaCl intake (4 mmol/kg/day) studied by other workers (Anderson et al., 2008). Suffice it to say that plasma [EO] cannot be used as an indicator of plasma volume in either normal or hypertensive subjects.
Despite lack of evidence for the natriuretic activity of EO, several lines of experimental evidence clearly demonstrate the prohypertensive role of EO, including induction of hypertension in ouabain-treated rodents, elevation of EO levels in hypertensive rats, and observations of the central prohypertensive action of this 1996a,b, 1998; Fedorova et al., 2005a, 2007). Increases in brain EO in Dahl-S rats are caused by central administration of NaCl or by NaCl loading; these increases are mediated by the brain RAS and by sympathoactivation (Huang and Leenen, 1996b). Leenen and coworkers demonstrated in rats that an increase in cerebrospinal fluid NaCl precedes the development of hypertension (Huang et al., 2004) and that sodium ions in the brain enter the intracellular space via epithelial sodium channels (Wang and Leenen, 2003). Moreover, they also demonstrated that this process is modulated by central mineralocorticoid receptors (Amin et al., 2005). Accordingly, Gabor and Leenen (2009) demonstrated in Wistar rats that immunoneutralization of brain EO prevented the potentiating effect of exogenously administered aldosterone on pressor response induced by central administration of sodium chloride. Some data, however, suggest that brain-specific rather than epithelial sodium channels are involved in this process (Orlov and Mongin, 2007).
Haupert and coworkers have described a specific pathway for the biosynthesis of EO in the hypothalamus of Milan hypertensive rats (Murrell et al., 2005). However, no such pathway could be identified in the adrenal cortex. Therefore, it seems that in NaCl-sensitive hypertension, EO is likely to act as a central regulator rather than as a peripheral effector. This concept is integral to the schema we propose in Fig. 4.
2. Endogenous Digoxin
one of the endogenous CTS represents digoxin. Thus, Goto et al. (1990) purified a substance indistinguishable from digoxin from human urine. Later, another group (Qazzaz et al., 1996) reported that bovine adrenal glands contain deglycosylated analogs of digoxin. It is noteworthy that digoxin (200 g/kg/day) has been previously reported to reverse hypertension induced by chronic ouabain administration (Huang et al., 1999). Although the concept of endogenous digoxin as an antagonist of EO in humans is quite appealing, the extensive variability of commercial digoxin immunoassays to detect endogenous CTS (Sheiner et al., 1974; Bergdahl et al., 1979; Bergdahl and Molin, 1981; Kelly, 1986; Goto et al., 1991a, 1992) argues strongly against this possibility.
B. Endogenous Bufadienolides
Amphibians are known to produce CTS of bufadienolide nature, which are different from cardenolides in having a doubly unsaturated six-membered lactone ring (Fig. 2) (Meyer and Linde, 1971). Bufadienolide-containing preparations from frog and toad skin have been (and still are) used for treatment of congestive heart failure (CHF) in the traditional medicine of the Far East (Chen and Kovaríková, 1967). It is noteworthy that the highest levels of bufadienolides are detected in the skin of those amphibian species, which migrate from a dry to an aquatic environment (Flier et al., 1980). Because in Amphibia the skin regulates water and electrolyte ho due to an increase in the resident time of the sodium pump in the cellular membrane (Efendiev et al., 2004; Ferrandi et al., 2004). In this strain of animals, ouabain apparently leads to further up-regulation of the renal Na+/K+-ATPase expression, renal sodium retention, and hypertension (Ferrandi et al., 2004). Administration of an ouabain antagonist, rostafuroxin (PST2238), a digitoxin derivative (Figs. 2 and 3), attenuates the hypertension in this strain (Ferrari et al., 1999).
Elevated levels of EO can also elevate blood pressure via inhibition of α2 Na+/K+-ATPase and promotion of Ca2+ entry via the Na+/Ca2+ exchanger in vascular smooth muscle (Zhang et al., 2005; Dostanic-Larson et al., 2006). Thus, the genetically engineered mice that express ouabain-resistant α2 Na+/K+-ATPase, unlike control mice with more ouabain-sensitive sodium pumps, do not manifest an increase in BP after chronic administration of ouabain (Dostanic-Larson et al., 2006). Accordingly, vascular smooth muscle from these mice with ouabain-insensitive α2 sodium pumps is insensitive to the pressor effect of ouabain (Dostanic-Larson et al., 2005). Furthermore, genetically engineered mice with reduced expression of α2 (but not of α1) Na+/K+-ATPase become hypertensive, and their arteries exhibit enhanced vascular tone in vitro (Zhang et al., 2005).
Alternatively, other workers have not found major differences in ouabain binding to the caveolar compared with noncaveolar Na+/K+-ATPase (Liang et al., 2007).
Liu et al. (2004, 2005) have observed that both signal transduction through the Na+/K+-ATPase-Src-EGFR cascade and clathrin-mediated endocytosis of the Na+/ K+-ATPase in renal proximal tubular epithelial cells are limited to the Na+/K+-ATPase residing in caveolae.
It is possible that different levels of expression of the γ subunit of the Na+/K+-ATPase in the preparations used explain some of these differences as suggested by Nguyen et al. (2007). In accord with either of these scenarios, patients with hypertension and mutations in the adducin gene also exhibit altered renal sodium reabsoption (Wang et al., 2003). Manunta et al. (2008) demonstrated that saline loading produces renal sodium retention in hypertensive patients with elevated plasma EO levels and mutant adducin. In Milan hypertensive rats, administration of the novel antihypertensive compound, PST2238 (Ferrari et al., 1999), antagonizes the interaction of EO and adducin on the renal sodium pump, lowers the BP, and inhibits the activity of Na+/ K+-ATPase in the renal medulla (Ferrandi et al., 2004).
In rats, centrally administered ouabain elicits pressor and natriuretic responses that depend on the activation of the renin-angiotensin system (RAS) (Takahashi et al., 1984; Yamada et al., 1994; Huang and Leenen, 1996a,b).
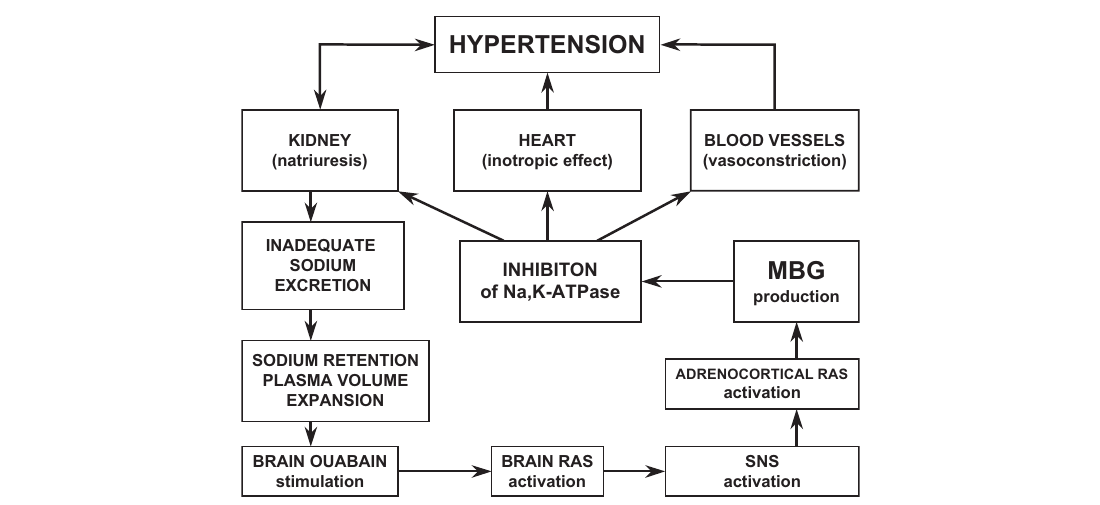
Likewise, there is substantial evidence to indicate that brain EO contributes to the pathogenesis of NaCl-sensitive hypertension and is stimulated by acute and chronic NaCl loading (Goto et al., 1992; Huang and Leenen, HYPERTENSION KIDNEY HEART (inotropic effect) BLOOD VESSELS INADEQUATE SODIUM EXCRETION INHIBITON of Na,K-ATPase production SODIUM RETENTION PLASMA VOLUME EXPANSION ADRENOCORTICAL RAS activation activation activation stimulation MBG
C. Biosynthesis of Cardiotonic Steroids
Despite substantial progress in the elucidation of the structure and mechanisms of action of CTS in mammals, the biosynthesis of these endogenous CTS is still poorly understood. Based on the hypothesis that CTS have a steroidal structure, Schreiber et al. (1981b), demonstrated that rat adrenal extracts contain a digitalis-like immunoreactive material. Since that time, evidence for the adrenocortical origin of CTS has grown. First, the de novo biosynthesis of EO was demonstrated in cultured adrenocortical cells (Perrin et al., 1997; Lichtstein et al., 1998; Qazzaz et al., 2004). el-Masri et al. (2002) found that human adrenocortical cells also produce dihydro-ouabain in addition to EO. In agreement with the classic scheme of adrenocortical steroidogenesis, Lichtstein et al. (1998) using radiolabeled hydroxycholesterol as a possible CTS precursor, demonstrated that the side chain cleavage of cholesterol is essential for synthesis of EO. The pharmacological inhibition of the next step of classic steroidogenesis (i.e., conversion of pregnenolone to progesterone) was shown to reduce the synthesis of EO by adrenocortical cells (Perrin et al., 1997). Unfortunately, the enzymes controlling the conversion of progesterone into CTS via inversion of the configuration at carbons 5 and 14 to form the cis-trans-cis configuration so far remain unknown (Hamlyn, 2004). Because we do not know the enzymes responsible for the synthesis of specific CTS, our ability to develop knockout and/or overexpression models has also been rather limited so far.
Murrell et al. (2005) found that the genes coding P450scc (an enzyme controlling the side chain cleavage of cholesterol) and ⌬5–3β-hydroxysteroid dehydrogenase isomerase (an enzyme that converts pregnenolone to progesterone) are up-regulated in the hypothalamus of Milan hypertensive rats but not in the hypothalamus of their normotensive counterparts (Milan normotensive rats). Accordingly, the knockdown of ⌬5–3β-hydroxysteroid dehydrogenase in rat brain led to a marked reduction in the production of sodium pump-inhibitory material (Murrell et al., 2005). Surprisingly, in the same study, no such mechanism of CTS biosynthesis was identified in the adrenal cortex of Milan hypertensive rats.
Evidence for neuroregulatory roles of EO and for the neuroendocrine source of EO has accumulated (Bagrov et al., 2002; Huang et al., 2006). Komiyama et al. (2001) specifically hypothesized that EO may be produced by and secreted from the adrenal medulla. These researchers purified a substance from the PC12 pheochromocytoma cell line that originated from progesterone and that, by its chromatographic and mass spectrometric properties, was not different from authentic ouabain (Komiyama et al., 2001). The above findings agree with a clinical observation of a marked reduction in the levels of circulating EO in a patient after the surgical removal of a pheochromocytoma (Komiyama et al., 1999).
In toads, bufadienolides are synthesized from cholesterol, as demonstrated by experiments with [14C]cholesterol (Siperstein et al., 1957). However, in several toad species, it has been demonstrated that pregnenolone is not incorporated into bufadienolides. This result indicates that cholesterol side chain cleavage is not involved in the biosynthesis of the predominant bufodienelide CTS in these species (Siperstein et al., 1957; Porto and Gros, 1970, 1971; Garraffo and Gros, 1986). Chen and Osuch (1969) demonstrated in toads that bufadienolide CTS may be synthesized from cholesterol via a “bile acid” pathway (i.e., from cholanates). A series of studies performed by Doris and coworkers confirmed that the biosynthesis of bufadienolide CTS does not involve the pathway implicated in synthesis of “classic” steroid hormones and EO. First, this group established that the important interaction between brain and peripheral CTS. It has been observed in both acute and chronic NaCl loading in Dahl-S rats that a transient increase in circulating EO precedes a sustained increase in circulating MBG (Fedorova et al., 2000, 2002b). This finding has led Fedorova et al. (2005a) to postulate that EO, acting as a neurohormone, triggers MBG production and secretion, which in turn effectuates the increases in cardiac contractility, peripheral vasoconstrictor, and natriuresis observed in this model (Fig. 4). Subsequently, Fedorova et al. (2007) demonstrated that in salt-loaded Dahl-S rats, the brain EO exhibits peak transient responses in the amygdala, hippocampus, and supraoptic nucleus of hypothalamus. From this study, brain EO seems to activate the central RAS, which, possibly via sympathoactivation, in turn activates the RAS in the adrenal cortex.
Activation of the adrenocortical RAS then seems to facilitate MBG production and secretion, which results in elevated plasma [MBG] and urinary MBG excretion.
This sequence of events (Fig. 4) was fully mimicked by intrahippocampal administration of a very low (60 pg) dose of plant-derived ouabain (Fedorova et al., 2007).
Komiyama et al. (2005), using tandem mass and NMR spectrometry, demonstrated that uremic plasma contains increased levels of another bufadienolide, telocinobufagin, in addition to MBG. Telocinobufagin is different from MBG in that it has a hydroxyl at the 14-position of the lactone ring versus an epoxy group in the 14,15-position. The authors hypothesized that telocinobufagin may be a natural precursor of MBG (Komiyama et al., 2005).
Bufalin is a very potent Na+/K+-ATPase inhibitor, and bufalin derivatives were the first bufadienolides positively identified in the mammals (Lichtstein et al., 1993). Bufalin-like immunoreactive material has been identified in human and rat tissues (Kieval et al., 1988; Goto et al., 1991a; Oda et al., 2001). Although levels of bufalin-like compound in plasma from hypertensive Dahl-S rats were found to be moderately elevated, little is known at present about the physiological significance of this CTS in mammals.
al., 1998; Sophocleous et al., 2003; Lorenz et al., 2008). It should be noted, however, that in a few clinical and experimental studies (Takahashi et al., 1988; Naruse et al., 1994; Bernini et al., 1998) levels of EO were found to be independent of adrenalectomy and ACTH administration. One explanation for these discrepancies is that, similar to other hormones (Miller, 2008), the biosynthesis of EO may occur both in the adrenal cortex and in extraadrenal tissues.
Because all classic steroid hormones circulate in a protein-bound form, it is not surprising that this also seems to be the case for mammalian CTS. Lichtstein et al. (1993) and Butler et al. (1996) addressed this issue.
They found that a substantial portion of bufadienolides are protein-bound in toads; in these animals, plasma levels of bufadienolides reach an enormously high (micromolar) level. Subsequently, Antolovic et al. (1998) isolated from mammalian plasma and kidneys a specific cardiac glycoside-binding globulin with higherand lower-affinity binding sites for ouabain and possibly for endogenous CTS. Later, CTS-binding protein was identified as a fragment of IgG (Komiyama et al., 1998; Parhami-Seren et al., 2002). The importance of CTSbinding protein(s) on the function of CTS has not been studied consistently yet; however, Antolovic et al. (2000) suggested that binding of CTS to and unbinding of CTS from this transport protein may be involved in a rapid and transient 36-fold rise of EO in response to exercise.
D. Differential Sensitivity of Na+/K+-ATPase Isoforms to Cardiotonic Steroids
to Cardiotonic Steroids A separate line of evidence indicates that various endogenous sodium pump ligands exhibit selectivity toward different isoforms of the Na+/K+-ATPase. Ferrandi et al. (1993) demonstrated that a Na+/K+-ATPase inhibitor purified from rat and bovine hypothalamus inhibits the Na+/K+-ATPase from rat brain (α3 isoform) similar to ouabain and surpasses ouabain in its ability to inhibit the renal sodium pump (α1 isoform). Another sodium pump inhibitor, a labile factor derived from the peritoneal dialysate of patients with chronic renal failure that is clearly dissimilar from ouabain in its elution profile, exhibited greater affinity toward the α1 and α2 rather than the α3 isoform of Na+/K+-ATPase (Tao et al., 1996, Graves et al., 2000). Subsequently, this material was shown to potently inhibit the Na+/K+-ATPase from rabbit kidney (α1 isoform) and by its vasoconstrictor activity exceeded ouabain by more than 2 orders of magnitude (Graves et al., 2000). Crambert et al. (1998) purified digitalis-like immunoreactive material from adult and neonatal plasma. This material exhibited greater affinity toward the α1 and α2 versus α3 Na+/ K+-ATPase isoforms.
Fedorova and Bagrov (1997) have studied mechanisms of the vasoconstrictor effects of ouabain and MBG in isolated rat aorta. In this preparation MBG preferentially inhibited the Na+/K+-ATPase from vascular production of digitalis-like immunoreactive material by adrenocortical cells did not depend on cholesterol side chain cleavage (Doris et al., 1989, 1994). Later, they demonstrated in murine Y1 adrenocortical cells that the de novo biosynthesis of MBG does require cholesterol as a precursor but does not involve the conversion of cholesterol to pregnenolone via side chain cleavage by P450scc as EO does (Dmitrieva et al., 2000, 2005). Lichtstein et al. (1998), using rat primary cultured adrenal cells and labeled precursors, demonstrated that production of bufalin-like immunoreactive material derives from pregnenolone and does require cholesterol side chain cleavage. Because in this study (Lichtstein et al., 1998) the detection of bufalin was based on an assay using antibody with very high (19.5%) cross-reactivity to the cardenolide strophanthidin, it is difficult to exclude the possibility that these results may, in fact, refer to the production of EO. Murine Y1 adrenocortical cells have been demonstrated to produce marinobufotoxin, an MBGsuberylarginine conjugate previously isolated from various amphibian tissues (Yoshika et al., 2007). At this juncture, it seems that although both bufadienelide and cardenolide CTS derive from cholesterol, only cardenolide CTS synthesis requires cholesterol side chain cleavage.
The release of endogenous CTS is controlled by various humoral stimuli including ACTH, ATII, vasopressin, and catecholamines. Release of EO from bovine adrenocortical cells is sensitive to ACTH, stimulation of α1adrenergic receptors agonists, and ATII (Laredo et al., 1994, 1995, 1997). In human adrenocortical cells, however, the release of EO is insensitive to ACTH and ATII, whereas it is stimulated by phenylephrine and vasopressin (Laredo et al., 1997). It is noteworthy that in bovine adrenocortical cells ATII acting via AT1 receptors stimulates the release of aldosterone, whereas the release of EO is regulated by AT2 receptors (Laredo et al., 1997). In rat adrenocortical cells, EO secretion was shown to increase after stimulation of nicotinic cholinoceptors (Göõz et al., 2004). el-Masri et al. (2002) demonstrated that, unlike for EO, production of endogenous dihydroouabain by human adrenocortical cells is stimulated by cAMP. In bovine adrenocortical cells, Shah et al. (1998) showed that AT2 receptor antagonists blocked the effects of ATII on release of EO but not aldosterone. Treatment of these cells with dibutyryl cAMP, on the contrary, stimulated secretion of aldosterone with no effect on release of EO. These mechanisms may explain in vivo rapid and dramatic peak responses of EO to stimuli such as exercise (Bauer et al., 2005) and acute NaCl loading (Fedorova et al., 2000). Fedorova et al. (2005), in primary cultures of rat adrenocortical cells, demonstrated that ATII via AT1 receptors stimulates the production of MBG.
Both clinical and experimental animal studies have shown that plasma levels of CTS, including EO and MBG, decrease after adrenalectomy and rise after administration of ACTH (Gault et al., 1988; Fedorova et smooth muscle sarcolemma (α1 isoform), whereas ouabain exhibited higher affinity toward Na+/K+ATPase from vascular nerve endings (α3 isoform). Subsequently, amphibian MBG and MBG-like immunoreactive material purified from hypertensive Dahl-S rats was found to be a potent inhibitor of ouabain-resistant Na+/K+-ATPase from rat kidney (Fedorova et al., 2000, 2001a).
Pierre et el. (2008), using ERK phosphorylation as an indicator of ouabain-induced signaling, demonstrated that α3/β1 and α4/β1 but not α2/β1 Na+/K+-ATPase responded to ouabain treatment, indicating the importance of isoform-specific differences in Na+/K+-ATPase signaling.
E. Hypertension and Cardiotonic Steroids
V. Endogenous Cardiotonic Steroids and Cell Signaling
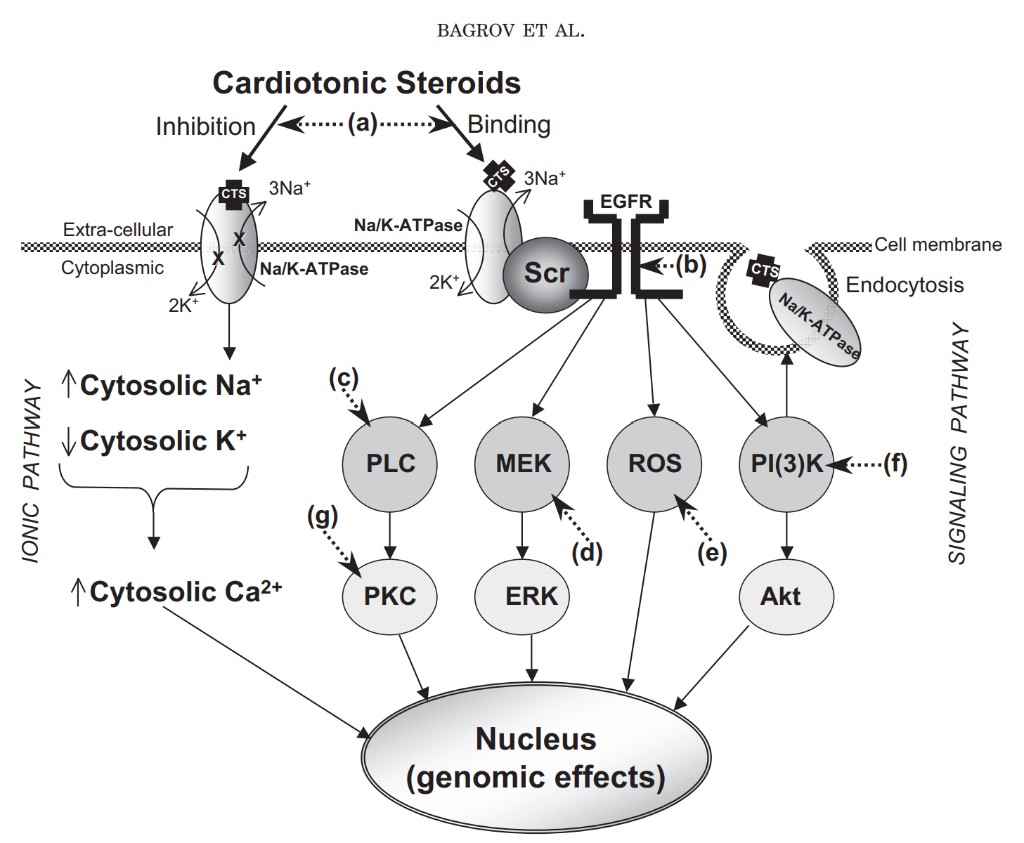
The first or classic model to explain interactions of endogenous CTS with the plasmalemmal Na+/K+-ATPase was based on the observation that many CTS inhibit the enzymatic and transport functions of the Na+/K+ATPase (Klein et al., 1971). The model, which has been enumerated in a large number of reviews, basically proposes that exposure of cells to CTS results in some degree of inhibition of the Na+/K+-ATPase pumping function and consequently an increase in cytosolic sodium, at least within some domain of the cell. This increase, in turn, alters the transport function of the Na+/Ca2+ exchanger, which then results in increases in cytosolic calcium, again, at least in some domain of the cell. With this connection to cytosolic [Ca2+], it is easy to see how innumerable cellular functions might be influenced (Blaustein, 1993). This model is illustrated in Inhibition 3Na+ CTS TS 3Na+ EGFR Na/K-ATPase X X C Na/K-ATPase Na /K -A TP as e Cytosolic K+ Cytosolic Ca2+ (genomic effects) Cytosolic Na+ Cell membrane CTS Binding
VI. Effects of Endogenous Cardiotonic Steroids on Kidney and Sodium Metabolism
As discussed in section III, a natriuretic substance is defined as one that increases urinary Na+ excretion; however, the prototypal CTS, digitalis or digoxin, is not natriuretic when administered in typical clinical doses in vivo to experimental animals or normal human subjects, although suprapharmacological doses of digoxin do alter sodium transport in the isolated kidney and impair transcellular transport of virtually all renal epithelial cells (Hauptman and Kelly, 1999). In contrast, administration of the atrial or brain natriuretic peptides induces prompt natriuresis in vivo and alters distal nephron sodium reabsorption in a well defined cGMP-mediated process (Itabashi et al., 1987; Nakamoto et al., 1987; Rubattu and Volpe, 2001; Lee and Burnett, 2007).
In distinction from digoxin or ouabain, the endogenous CTS described by Hillyard et al. (1976) did have an appreciable effect on renal Na+/K+-ATPase activity.
Work from our laboratories has demonstrated that MBG has potent effects on rat renal Na+/K+-ATPase activity, which can be reversed in vivo by administration of antiMBG antibodies. In addition to the direct inhibition of the Na+/K+-ATPase by MBG, other studies from our laboratories have shed new light on this subject. We observed that the administration of CTS induced endocytosis of the plasmalemmal Na+/K+-ATPase in LLCPK1 cells, a cell line maintaining features of proximal tubule cells (Liu et al., 2002). In fact, CTS actually induced decreases in plasmalemmal Na+/K+-ATPase density in a doseand time-dependent manner in these cells, whereas in MDCK cells, a cell line resembling distal tubular cells, no such depletion of the plasmalemmal Na+/K+-ATPase was observed. Among the CTS that were studied in LLC-PK1 cells, digoxin, MBG, and ouabain differed in their ability to induce this endocytosis; specifically, digoxin seemed to be the least effective whereas other CTS (e.g., MBG) were substantially more effective, even when results were normalized for the ability of the CTS to inhibit the Na+/K+-ATPase enzymatic function. Further studies demonstrated that CTS could actually inhibit transcellular Na+ transport in LLC-PK1 cells (Liu et al., 2002). Endocytosis of the Na+/K+-ATPase was subsequently shown to proceed through clathrin-coated pits and require PI(3)K activation and the plasmalemmal pump being in the context of caveola as well as signaling through the Src-EGFR pathway (Liu et al., 2004, 2005).
Extending these studies to an in vivo rat model, we observed evidence for CTS (specifically MBG)-induced endocytosis participating in the altered sodium reabsorption seen with increases in dietary sodium. Specifically, we observed that increases in dietary sodium induced increased urine excretion of MBG along with decreased proximal tubular Na+/K+-ATPase expression along with increased presence of the Na+/K+-ATPase in both early and late endosomes. Administration of an antibody to MBG blocked both the endocytosis of the Na+/K+-ATPase and blunted the increase in urinary sodium excretion seen with the high-salt diet (Periyasamy et al., 2005). Further work demonstrated that CTS can induce decreases in the apical expression of one of the plasma membrane Na+/H+ exchangers, NHE3 (Oweis et al., 2006; Liu and Shapiro, 2007). Some of the long-term decrease in NHE3 expression is related to decreases in NHE3 transcription, whereas, acutely, binding of ouabain to the basolateral Na+/K+-ATPase seems to rapidly induce endocytosis of the apical NHE3 (Cai et al., 2008). Taken together, these data suggest that increases in the circulating levels of MBG accompany salt loading, which may in turn induce decreases in both basolateral and apical sodium transport in the proximal tubule through both the classic (ionic) or signaling mechanism and the more recently described Na+/ K+-ATPase-Src-EGFR pathway. This decrease in renal sodium reabsorption would then result in increases in urinary sodium excretion (Fig. 5). Therefore, through direct effects on renal epithelium, some CTS do seem to function as natriuretic substances in vitro and in vivo (Liu and Shapiro, 2007).
In addition to the effects of bufadienolides on the heart and vascular system, these hormones have renal al., 1991; Numazawa et al., 1994; Kawazoe et al., 1999; Ghoumari et al., 2006; Larre et al., 2006; RodriguesMascarenhas et al., 2006).
The debate between the classic (ionic) and alternative (signaling) pathways may ultimately end in a tie. Although it seems clear that the alternative pathway can explain many of the effects of CTS, it does not seem to explain all of them. For example, it is well known that red blood cells of patients with end-stage renal disease have decreases in ouabain-sensitive rubidium uptake and enzymatic Na+/K+-ATPase function that can be reversed by incubation, in vitro, with antibodies to digitalis and/or other CTS (Periyasamy et al., 2001; Bagrov et al., 2005). Another point is that among Na+/K+-ATPase isoforms, predominantly the α1 isoform seems to reside in caveolae in close physical proximity to Src.
However, it is very clear from the elegant studies of Lingrel and colleagues that changes in the CTS sensitivity of the α2 isoform produce profound physiological effects (Dostanic et al., 2003, 2004, 2005). In addition, Feldmann et al. (2007) have demonstrated that even the endocytosed Na+/K+-ATPase may still have a pumping function; these investigators have been able to distinguish the pH within early endosomes, depending on whether ouabain still interacted with the endocytosed Na+/K+-ATPase. From these and other data that have been summarized in several reviews, we propose that the classic and alternative pathways may work both in parallel and synergistically to effect physiological consequences of CTS binding to the Na+/K+-ATPase.
VII. Chronic Renal Failure and Uremic Cardiomyopathy
The modern concept of uremic cardiomyopathy is that patients with renal disease develop a progressive loss of diastolic function and ventricular hypertrophy, which ultimately over time may progress to eccentric hypertrophy and, rarely, systolic dysfunction (Mohmand et al., 2005). Based on echocardiographic studies, diastolic dysfunction and ventricular hypertrophy are extremely common whereas systolic dysfunction occurs in less than 20% of patients with end-stage renal disease (ESRD); neither the diastolic dysfunction nor the ventricular hypertrophy can be explained only by the hypertension and anemia that usually complicate ESRD (Middleton et al., 2001). As mentioned in section IV.B, plasma levels of telocinobufagin and MBG have been noted to be substantially elevated in patients with ESRD (Komiyama et al., 2005). To examine this important issue, our laboratory has established a model of chronic renal failure using partial nephrectomy in the rat (Kennedy et al., 2003) and the mouse (Kennedy et al., 2007). Animals subjected to partial nephrectomy develop increases in MBG similar to that seen clinically in subjects with renal failure (Gonick et al., 1998; Kennedy et al., 2006a); these animals also develop other features similar to human uremic cardiomyopathy including diastolic dysfunction and ventricular hypertrophy as well as evidence for signaling through the Na+/K+-ATPase-SrcEGFR cascade and ERK activation (Kennedy et al., 2003, 2007). In this rodent model, cardiac fibrosis is especially prominent (Kennedy et al., 2006a, 2007; Elkareh et al., 2007), although to be fair, it also seems to complicate human uremic cardiomyopathy (Harnett and Parfrey, 1994; London and Parfrey, 1997). In the rat, infusion of MBG to achieve elevations in plasma [MBG] similar to those seen with partial nephrectomy results in activation of the Na+/K+-ATPase-Src-EGFR cascade and ERK and many of the phenotypical features of experimental uremic cardiomyopathy, whereas active immunization against MBG attenuates most of the biochemical, physiological, and morphological features of uremic cardiomyopathy in animals subjected to partial nephrectomy (Kennedy et al., 2006a; Elkareh et al., 2007).
Additional work in this area has demonstrated that in the rodent very small amounts of MBG (and other CTS), virtually identical to the circulating plasma concentrations seen in experimental and clinical renal failure, directly stimulate the production of collagen in the primary culture of cardiac fibroblasts (Elkareh et al., 2007).
Again, this direct effect seems to require signaling through the Na+/K+-ATPase-Src-EGFR cascade. Although some activity of the transforming growth factorβ-Smad pathway is necessary for this stimulation, no changes in the amount of transforming growth factor-β or the Smads were identified. The stimulation of collagen production was associated with increased translational procollagen as well as increases in mRNA for procollagen, whereas no change in procollagen or collagen stability could be identified (Elkareh et al., 2007).
Low concentrations of CTS have been shown to induce collagen synthesis by dermal fibroblasts, which could potentially be exploited to accelerate wound healing (ElOkdi et al., 2008).
VIII. Congestive Heart Failure
CHF is associated with fluid retention and plasma volume expansion; therefore, one would expect CTS to be stimulated under these circumstances (Schrier and Abraham, 1999). As early as 1981, Schreiber et al.
(1981a) hypothesized that CTS might be involved in the regulation of tissue growth and myocardial hypertrophy.
Morise et al. (1988) demonstrated that development of CHF in rats is associated with increased plasma Na+/ K+-ATPase inhibitory activity. Liu et al. (1990) showed in a group of 50 patients that the severity of CHF is positively associated with plasma levels of digoxin-like immunoreactivity as well as erythrocyte sodium concentrations. The relationship between CTS, cardiac geometry, and central hemodynamic parameters has been analyzed in several studies. Gottlieb et al. (1992) found that although plasma EO did not exhibit graded increases with the progression of cardiac failure, EO levels were elevated in patients with severely impaired left ventricular (LV) performance (LV ejection fraction less than 21%). Manunta et al. (1999) demonstrated that plasma EO positively correlated with systolic and diastolic blood pressure in a group of 110 normotensive subjects and 100 hypertensive subjects; these workers found that EO levels positively correlated with LV mass effects that are important physiologically. First, increases in MBG accompany decreases in renal function; this has been established both in experimental animals and humans (Gonick et al., 1998; Priyadarshi et al., 2003; Komiyama et al., 2005; Kennedy et al., 2006a, 2007). More importantly, it has been demonstrated that MBG induces endocytosis of the proximal tubular Na+/ K+-ATPase and decreases renal sodium reabsorption, both in vitro and in vivo; in particular, our group has demonstrated that administration of antibodies to MBG alters the endocytosis of the proximal tubular Na+/K+ATPase and decreases urinary sodium excretion in Sprague-Dawley rats (Liu et al., 2002; Periyasamy et al., 2005). In agreement with the notion that inhibition of renal α1 Na+/K+-ATPase by MBG represents one of the mechanisms regulating renal sodium excretion, genetically engineered mice that express ouabain-sensitive α1 Na+/K+-ATPase have been shown to exhibit greater natriuretic responses to saline loading than mice with ouabain-insensitive α1 Na+/K+-ATPase (Loreaux et al., 2008).
the cardiac Na+/K+-ATPase to ouabain (Fedorova et al., 2004).
It has been demonstrated that ouabain and MBG in vitro in cultured renal tubular cells induce apoptosis and growth-promoting signaling, respectively (Akimova et al., 2005). The existence of endogenous CTS with distinct effects on cell survival makes teleological “sense,” because different in vivo scenarios may require modulation of sodium pump-dependent functions with or without cytotoxic actions. Thus, the transition from compensated LV hypertrophy to CHF is accompanied by a decrease in the plasma MBG and by a 3-fold increase in plasma EO when the induction of cell death may be desirable (Neuss et al., 2001).
Involvement of CTS in pathogenesis of experimental CHF is not limited to hypertensive models. In rats with CHF complicating experimental myocardial infarction, the chronic blockade of brain EO with Digibind (Fab fragments of polyclonal anti-digoxin antibody developed for treatment of digoxin intoxication) substantially inhibited the development of LV dilation and dysfunction (Leenen et al., 1999).
In humans, the development of CHF is associated with decreased Na+/K+-ATPase activity in LV myocardium along with a decline in the expression of both α1 and α3 Na+/K+-ATPase isoforms (Schwinger et al., 1999). In animal studies, changes in Na+/K+-ATPase isoform composition observed in the context of LV remodeling are complex and seem to be specific to both the species and experimental models. Thus, similar to those observed in hypertensive Dahl-S rats (Fedorova et al., 2004), significantly elevated LV levels of α1 Na+/K+ATPase have been reported in rats with diabetic cardiomyopathy (Gerbi et al., 1997). In other studies of LV hypertrophy occurring in hypertensive rats, levels of α1 Na+/K+-ATPase in LV myocardium were unchanged, whereas those of α2 Na+/K+-ATPase were decreased (Book et al., 1994, Charlemagne et al., 1994).
In short, clinical and experimental studies demonstrate that in CHF levels of CTS change and accompany changes in the cardiac expression of their molecular target, the Na+/K+-ATPase. Considering growing evidence for an important role of CTS in cell signaling, growth, and apoptosis, it seems clear that these molecules represent potential biomarkers for CHF and cardiovascular remodeling as well as potential therapeutic targets.
IX. Preeclampsia
Despite the fact that preeclampsia occurs in between 3 and 5% of all pregnant women and represents the leading cause of maternal and fetal mortality, its pathogenesis remains poorly understood. Moreover, treatment for preeclampsia has not changed significantly in many years (Funai et al., 2005; Sibai et al., 2005). That said, index and LV end diastolic volume in only the hypertensive subjects. Later, Pierdomenico et al. (2001) found that circulating EO levels in 92 hypertensive patients were positively correlated with mean BP and total peripheral resistance index, whereas LV end-diastolic volume index, stroke index, and cardiac index exhibited inverse correlations with the plasma EO. Plasma EO was found to independently predict total peripheral resistance index, cardiac index, and relative wall thickness. Moreover, the plasma EO was substantially higher in patients with eccentric remodeling compared with those subjects with normal LV geometry or concentric hypertrophy (Pierdomenico et al., 2001). In another study performed in patients with LV dysfunction (Balzan et al., 2001), the plasma [EO] was found to be elevated compared with that in normal subjects but did not correlate with the LV ejection fraction. Fridman et al.
(2002) studied 23 consecutive hypertensive male patients with CHF; in these patients, the plasma MBG exhibited progressive increases that paralleled the progression of CHF and increases in plasma α-human atrial natriuretic peptide (α-hANP). Although in the same study, the plasma EO was not found to be elevated compared with that in normal subjects, the levels of EO were found to be substantially elevated in a subset of patients with CHF who had ejection fractions less than 30% (A. Y. Bagrov and V. I. Novikov, unpublished observations), similar to the observation of Gottlieb et al.
Experimental data also indicate an association between elevated plasma CTS and cardiovascular remodeling. Normotensive rats consuming a 4% NaCl diet demonstrate increases in plasma MBG and correlated increases in cardiac weight in the absence of hypertension (Fedorova et al., 2001a). Sustained ouabain infusion, which causes a 2-fold elevation of plasma ouabain immunoreactivity, is also sufficient to induce LV hypertrophy in normotensive rats (Ferrandi et al., 2004).
Studies performed in Dahl-S rats subjected to a highsalt diet coordinated shifts between the physiological function of the left ventricle (first compensated LV hypertrophy develops, which later transitions into a dilated cardiomyopathy), plasma levels of CTS, and the amounts of α1 and α2 Na+/K+-ATPase isoforms within the LV myocardium (Fedorova et al., 2004). Specifically, the advancing stages of hypertrophy were associated with elevated plasma MBG, increased levels of α1 (compared with α2) Na+/K+-ATPase in the LV myocardium, and a heightened sensitivity of the cardiac sodium pump to MBG. The transition to CHF was associated with a decline in MBG levels and decreased levels of α1 Na+/ K+-ATPase protein in the left ventricle. Levels of EO in Dahl-S rats with CHF rose substantially, and the enhanced EO production was associated with increased levels of ouabain-sensitive α3 Na+/K+-ATPase in LV myocardium, along with an increase in the sensitivity of caused a substantial increase in the activity of the sodium pump in thoracic aortae. A similar pattern of CTS levels has been observed in another model of preeclampsia, specifically, pregnant rats treated with deoxycorticosterone acetate and given 0.9% saline as drinking water for the duration of their pregnancy (Vu et al., 2005). Uterine arteries from these rats exhibited enhanced sensitivity to the vasoconstrictor effects of MBG.
Moreover, anti-MBG antibody substantially reduced the blood pressure in these animals. It has been demonstrated that in patients with preeclampsia, a monoclonal antibody against MBG detects elevated levels of MBG in plasma and placenta and ex vivo restores activity of the Na+/K+-ATPase in erythrocytes (Fedorova et al., 2008).
In preeclampsia, besides exhibiting direct vasoconstrictor effects, heightened CTS levels can alter the function of cytotrophoblasts and thus affect placentation. In cultured human cytotrophoblast cells, MBG, via activation of c-Jun NH2-terminal kinase, p38, and Src and possibly via induction of apoptosis, has been shown to impair cell proliferation, migration, and invasion (Uddin et al., 2008a,b).
Thus, it seems that CTS contribute to pathogenesis of preeclampsia and that increases in [MBG] rather than changes in [EO] are responsible for preeclampsia-induced Na+/K+-ATPase inhibition. This notion is consistent with the bufadienolide nature of endogenous CTS purified from human placentae (Hilton et al., 1996).
X. Other Conditions
Considering the fact that the CTS are one of the factors regulating transport and signaling functions of a key membrane enzyme, Na+/K+-ATPase, it is not a surprise that the list of disorders in which CTS are implicated, is not limited to essential hypertension, ESRD, preeclampsia, and cardiac failure.
A. Myocardial Ischemia/Infarction
Arrhythmogenicity is one of the most serious side effects of digitalis glycosides, and acute myocardial ischemia is known to be associated with heightened sensitivity of LV myocardium to the arrhythmogenic action of digitalis (Akera and Brody, 1977). Maixent and Lelièvre (1987) demonstrated that acute myocardial ischemia is associated with a loss in the number of high-affinity ouabain binding sites and with inhibition of the Na+/ K+-ATPase in canine LV myocardium. Bagrov et al.
(1989) hypothesized that endogenous CTS may be responsible for myocardial ischemia-induced inhibition of LV Na+/K+-ATPase and that heightened CTS levels may contribute to ischemia-induced ventricular arrhythmias. In agreement with this notion, Delva et al.
(1990) observed a rapid increase in plasma Na+/K+ATPase inhibitory activity in patients with coronary artery disease associated with decreased cardiac output during a transient period of myocardial ischemia while one theory of the pathogenesis of preeclampsia attributes an important role to CTS.
Because pregnancy is associated with plasma volume expansion involving renal sodium and fluid retention (Gallery et al., 1979; Masilamani et al., 1994), it is logical to examine the role of CTS in pregnancy and pregnancy-associated diseases. Buckalew and coworkers (Gusdon et al., 1984) were the first to demonstrate increased circulating levels of CTS in pregnancy and to hypothesize that CTS are involved in the pathogenesis of pregnancy-induced hypertension and preeclampsia.
Subsequently, several groups of investigators demonstrated elevated levels of digitalis-like immunoreactivity and elevated Na+/K+-ATPase inhibitory activity in preeclamptic plasma (Graves et al., 1984; Graves, 1987; Shrivastav et al., 1988; Poston et al., 1989; Kaminski and Rechberger, 1991; Gilson et al., 1997). After establishment of specific assays for cardenolide and bufadienolide CTS, levels of both EO and MBG were found to be moderately increased in normotensive pregnant women. In contrast, MBG but not EO was markedly elevated in patients with severe preeclampsia (average blood pressure 160/104 mm Hg) (Lopatin et al., 1999). In subjects with milder preeclampsia (blood pressure 149/93 mm Hg), elevated levels of MBG but, again, not of EO were observed and accompanied the observed inhibition of Na+/K+-ATPase in the erythrocytes. Ex vivo treatment of erythrocytes derived from these preeclamptic women with anti-MBG, but not anti-ouabain antibody, reversed the preeclampsia-associated Na+/K+-ATPase inhibition (Averina et al., 2006).
Although the exact role(s) of CTS in pregnancy is not yet understood, Digibind lowered the blood pressure in 15 patients with preeclampsia (Goodlin, 1988; Adair et al., 1996, 2009). The mechanism underlying this effect is believed to be cross-reactivity with endogenous CTS.
These clinical observations are in agreement with the results of an experimental study, which demonstrated a vasorelaxant action of Digibind in isolated perfused preeclamptic placentae (Di Grande et al., 1993). Accordingly, the Na+/K+-ATPase from preeclamptic placentae was shown to be up-regulated and to exhibit heightened sensitivity to digitalis glycosides (Amler et al., 1994).
This concept has been explored further in experimental animals. In pregnant Sprague-Dawley rats, levels of both MBG and EO doubled by the end of gestation.
Sodium chloride (1.8%) supplementation of rats during days 14 to 20 of gestation was associated with a 33 mm Hg increase in systolic blood pressure and proteinuria and decreases in fetal weight, size, and number of offspring (Fedorova et al., 2005b). Compared with normotensive pregnant rats, the development of hypertension in NaCl-supplemented pregnant rats was accompanied by increased plasma [MBG] but not [EO] (Fedorova et al., 2005b). In the same study, administration of antiMBG antibody to NaCl-supplemented pregnant rats resulted in a 28 mm Hg decrease in blood pressure and undergoing percutaneous coronary angioplasty. Subsequently, it was demonstrated in patients with acute myocardial infarction that an increase in plasma digoxin-like immunoreactivity accompanies the inhibition of erythrocyte Na+/K+-ATPase (Bagrov et al., 1991, 1994).
Accordingly, pretreatment with anti-digoxin antibody was shown to prevent myocardial ischemia-induced inhibition of cardiac Na+/K+-ATPase and to decrease the incidence of ventricular arrhythmias in coronary arteryligated rats (Bagrov et al., 1993b; Ke et al., 2004). Subsequently, Bagrov et al. (1998) demonstrated that an elevation in plasma CTS could be attributed to increases in plasma MBG and purified MBG from urine of patients with myocardial infarction.
B. Diabetes Mellitus
C. Obstructive Sleep Apnea
Obstructive sleep apnea (OSA) is frequently associated with the metabolic syndrome, a condition that, in turn, is characterized by associated volume-dependent hypertension (Ehlenz et al., 1991) and impaired glucose tolerance (Levinson et al., 1993; Coughlin et al., 2004).
In OSA, repeated episodes of apnea/hypopnea lead to an increase in the arterial tension of carbon dioxide (PaCO2) (Fuse et al., 1999). Elevated PaCO2 levels decrease plasma pH and produce renal sodium retention, a major stimulus for CTS production (Anderson et al., 1995; Fedorova et al., 1996). Bagrov et al. (1995) observed that in healthy humans, the pressor response to voluntary hypoventilation is associated with a substantial inhibition of Na+/K+-ATPase in the erythrocytes and with a simultaneous increase in plasma levels of MBG but not of EO. In agreement with this observation, Paci et al. (2000) demonstrated that hypertensive subjects with OSA exhibit elevated plasma levels of an unidentified endogenous Na+/K+-ATPase inhibitor, which, by its chromatographic behavior, is different from EO and resembles MBG. In 52 patients with OSA, Zvartau et al. (2006) demonstrated that levels of MBG were elevated in proportion to the severity of the OSA.
In the same study, in patients with OSA 24-h MBG excretion exhibited positive correlations with diastolic blood pressure and plasma levels of leptin and insulin.
This finding suggests that in these patients, MBG may be one of the factors linking mechanisms of the pathogenesis of hypertension and insulin resistance (Zvartau et al., 2006).
D. Exercise
Early evidence indicated that prolonged strenuous exercise increased the circulating levels of CTS (Clerico et al., 1988), whereas decreased levels of digoxin-like immunoreactive CTS were implicated in the blood pressure-lowering effects of the exercise (Koga et al., 1992; Komiyama et al., 1997). Bauer et al. (2005) reported intriguing data demonstrating that within 15 min of ergometry, athletes demonstrated 30-fold increases in the plasma levels of EO that rapidly decreased (within 5 min) with rest. A similar observation was made in beagles exposed to moderate exercise (Bauer et al., 2005).
These observations demonstrate that very dramatic effects of EO may be transient. This finding seems to be in agreement with reports demonstrating that EO can be implicated in behavioral stress (Goto et al., 1995, Weidemann et al., 2004) and via activation of hypothalamicpituitary-adrenocortical axis contributes to the onset of NaCl-sensitive hypertension, which was discussed previously (Fedorova et al., 2005a).
E. Behavioral Stress
Several observations demonstrate possible involvement of endogenous ouabain-like compound in behavioral stress. Goto et al. (1995) demonstrated in rats that swimming stress is associated with enhanced levels of ouabain-like immunoreactivity in plasma and adrenal glands but not in the pituitary gland and hypothalamus.
Fedorova et al. (2001a) have shown that social isolation of rats is associated with a transient increase in the renal excretion of ouabain-like material.
Elevated levels of CTS, along with perturbed functions of the Na+/K+-ATPase, were found in patients and experimental animals with diabetes mellitus (Clerico and Giampietro, 1990; Chen et al., 1993; Straub et al., 1996). Because insulin resistance often accompanies NaCl-sensitive hypertension, it has been hypothesized that in type 2 diabetes, CTS rise as a result of insulindependent renal sodium retention and that excessive CTS elaboration contributes to hypertension (Chen et al., 1993; Martinka et al., 1997). Additional evidence indicates that rats with type 1 diabetes exhibit greater levels of MBG and more profound sodium pump inhibition than rats with type 2 diabetes (Bagrov et al., 2005).
Along with clinical data demonstrating that CTS become stimulated during an oral glucose challenge (Carroll et al., 2001), this evidence indicated that CTS may also be implicated in determining insulin sensitivity. Accordingly, our laboratory demonstrated that in vivo administration of anti-MBG antibodies to rats after an oral glucose challenge produced greater elevations of plasma glucose and insulin compared with that in the vehicletreated animals (Bagrov et al., 2007). Kotova et al.
(2006) demonstrated that ouabain and MBG increase glycogen synthesis in skeletal muscle. This effect was additive to insulin and was mediated by activation of a Src-, ERK1/2-, p90rsk-, and glycogen synthase kinase 3-dependent signaling pathway. In skeletal muscle cells, MBG exceeded ouabain both with respect to sodium pump inhibition and induction of cell signaling. Thus, physiological mechanisms of regulation of carbohydrate metabolism and of glucose tolerance may involve CTS.
F. Mood Disorders
G. Ethanol Addiction
Neurochemical events associated with the effects of ethanol and with the development of ethanol addiction include changes in Na+/K+-ATPase activity. The effects of ethanol may be associated with both inhibition and activation of the sodium pump, depending on the dose, route of administration, and length of exposure (Nhamburo et al., 1986, 1987; Foster et al., 1989; Foley and Linnoila, 1995). Foley and Rhoads (1994) demonstrated a stimulatory effect of ethanol on the Na+/K+-ATPase in rat cerebral cortex synaptosomes; this is apparently related to its action on ouabain-sensitive (α2 and α3) rather than relatively ouabain-resistant (α1) Na+/K+-
H. Cancer
One of the most novel areas is the current CTS research examining the pathogenesis and treatment of cancer. Here, the initial evidence came from the clinical studies demonstrating that patients with breast cancer who were taking digitalis exhibited less recurrence and better survival than subjects not receiving digitalis treatment (Stenkvist et al., 1982; Mijatovic et al., 2007).
This rather staggering observation, taken together with a mounting body of evidence of the growth-promoting but anti-cancer effects of CTS in vitro, suggests the possibility of using digitalis drugs to treat cancer (Mijatovic et al., 2007). It is noteworthy that CTS may also be involved in cancer development. In a group of 84 patients with breast cancer, 73.6% had plasma levels of digitalis-immunoreactive CTS that were 3 times lower than those seen in control subjects, whereas 10.8% of these patients with cancer exhibited extremely high CTS levels (Weidemann et al., 2004; Weidemann, 2005).
Therapeutic strategies involving either addition of or antagonism of CTS are currently under development (Mijatovic et al., 2007).
In many studies both mania and bipolar depression have been associated with decrements in red blood cell Na+/K+-ATPase activity (Naylor et al., 1973; Choi et al., 1977; Hesketh et al., 1977; Rybakowski and Lehmann, 1994). Subsequently, Grider et al. (1999) studied plasma levels of digoxin-like immunoreactive factor in manic bipolar patients. In this study, normal subjects exhibited seasonal variations in plasma digoxin-like factor; levels of this compound were significantly lower in winter than those in spring, summer, and autumn. Manic subjects did not exhibit such seasonal variations, the level of digoxin-like immunoreactivity in winter was comparable with that in healthy control subjects, and no changes were observed in ill subjects during spring, winter, and autumn (Grider et al., 1999). Goldstein et al. (2006) demonstrated that levels of ouabain-like immunoreactivity in the parietal cortex of bipolar patients are significantly higher than those in normal individuals and depressed patients. These observations suggest that the defect in the Na+/K+-ATPase could be an important factor in pathogenesis of mood disorders (el-Mallakh and Wyatt, 1995).
Experimental findings support this contention. Acute intracerebroventricular administration of 5 l of 1 mM ouabain solution in rats produced a “mania-like hyperactivation” of the animals studied with automated activity monitors (el-Mallakh et al., 1995). Later, the same group reported that persistent hyperreactivity induced by central administration of ouabain was associated with substantial increases in ouabain binding and Na+/K+-ATPase activity in the hippocampus (Ruktanonchai et al., 1998). Pretreatment of animals with lithium prevented the above effects of ouabain (Li et al., 1997). There is additional evidence that Na+/K+ATPase inhibitors may affect mood and behavior; several amphibian-derived preparations containing bufadienolides have been used as recreational drugs with apparently addictive consequences (e.g., “toad licking psychosis”) (Howard and Foerstl, 1990; Lyttle et al., 1996).
ATPase isoforms. These authors concluded that ethanol induces sodium pump activation possibly via antagonizing the effect of an unidentified Na+/K+-ATPase endogenous inhibitory factor (Foley and Rhoads, 1994). Considering the involvement of Na+/K+-ATPase in the effects of ethanol and the existence of multiple endogenous CTS in the mammalian brain, Bagrov et al. (1996c) hypothesized that CTS may be implicated in the development of alcohol addiction. In rats, administration of digoxin (chosen to mimic the action of CTS) caused a decrease in voluntary alcohol consumption whereas immunization of the animals against digoxin, ouabain, and MBG markedly potentiated voluntary ethanol consumption (Bagrov et al., 1996c, 1999). In Wistar rats, ethanol administration stimulated cerebrospinal EO and peripheral MBG (Bagrov et al., 1999). In addition, MBG apparently mediates the pressor response to ethanol withdrawal in rats, which is associated with renal sodium retention (Kashkin et al., 2008). Therefore, in rats, ethanol administration stimulates CTS and suppresses the free choice of alcohol, whereas immunization against MBG and ouabain is associated with alcohol-seeking behavior. Based on the similarities in the neurobiology of depression and drug/alcohol dependence, Markou et al. (1998) suggested that the development of dependence may reflect self-medication of substances of abuse (i.e., alcohol) to normalize altered neurobiological mechanisms. Thus, the deficit of endogenous CTS might be expected to cause depression-like disorders and facilitate self-medicating behaviors, and CTS may be one of the factors linking the pathogenesis of mood disorders and ethanol dependence.
XI. Possible Strategies for Pharmacological Antagonism of Endogenous Cardiotonic Steroids
A. Blockade of Receptor Site on the Na+/K+-ATPase
B. Immunoneutralization
Although at first glance the in vivo immunoneutralization of CTS may appear to be an eccentric approach to the treatment of hypertension and blockade of volumesensitive hormones in a volume-contracted state may seem counterintuitive, clinical evidence for the efficacy of Digibind in preeclampsia comprises one of the most convincing arguments in favor of the prohypertensive effects of CTS. Goodlin (1988) reported a decrease in blood pressure in a 25.5-week preeclamptic patient after two intravenous infusions of Digibind (0.087 mg/kg each). Later, Adair et al. (1996) reported another case of successful use of Digibind in preeclampsia. Subsequently, the same group, in a placebo-controlled doubleblind study, demonstrated that Digibind (0.76 mg/kg) lowered the blood pressure in 13 patients with postpartum preeclampsia (Adair et al., 1996, 2009). It is noteworthy that Digibind did not exert adverse effects in these studies. It is hoped that the results of a multicenter, double-blind, placebo-controlled study of the efficacy of Digibind in preeclampsia (http://clinicaltrials.
gov/show/NCT00158743) will elucidate the potential utility of CTS immunoneutralization in preeclampsia.
The preliminary results of this study indicate that administration of Digibind to patients with severe preeclampsia is associated with an improvement in renal function manifested by an increase in creatinine clearance and by a reduction of plasma Na+/K+-ATPase inhibitory activity (Hopate et al., 2008; Lam et al., 2008).
The clinical impact of CTS immunoneutralization may not be limited to hypertensive emergencies. In fact, CTS antagonism may also be effective in patients with cerebral salt-wasting syndrome, a condition frequently accompanying cerebral injury and associated with a lifethreatening natriuresis. Digibind was shown to reduce renal sodium excretion in a patient after brain tumor removal (Menezes et al., 2003). Likewise, the magnitude of the pressor response after intracerebroventricular infusion of blood correlated with plasma sodium pumpinhibitory activity, and Digibind in vivo reduced natriuresis in rats.
C. Modulation of Interaction of Cardiotonic Steroids with Na+/K+-ATPase
with Na+/K+-ATPase 1. Protein Kinase C. Protein kinases are among the factors that regulate Na+/K+-ATPase phosphorylation, activity, and sensitivity to CTS (Blanco et al., 1998). In particular, PKC and PKG can affect the Na+/K+-ATPase activity and sensitivity to CTS via altering its phosphorylation state (Feschenko and Sweadner, 1997; Vasilets et al., 1999). The modulatory effect of PKC is specific to the Na+/K+-ATPase α1 isoform (Blanco et al., 1998; Fedorova et al., 2002a). In human arterial sarcolemma, PKC induces the phosphorylation of α1 Na+/K+ATPase and enhances the sensitivity of Na+/K+-ATPase to nanomolar concentrations of MBG (Fedorova et al., 2002a). Accordingly, in isolated human mesenteric arteries, nonselective activation of PKC by phorbol diesters markedly potentiates vasoconstrictor activity of MBG (Fedorova et al., 2002a). This synergistic interaction of MBG and PKC on vascular Na+/K+-ATPase is likely to participate in fine-tuning of the sodium pump The potential roles of spironolactone and canrenone (the major metabolite of spironolactone) as CTS antagonists (Semplicini et al., 1995) may be very interesting.
Excellent experimental studies demonstrate that both spironolactone and canrenone, which acts as a partial agonist of the digitalis receptor site on the Na+/K+ATPase, antagonize ouabain binding as well as shift Na+/K+-ATPase inhibition by ouabain (Finotti and Palatini, 1981; Garay et al., 1985). Canrenone has even been proposed as a treatment for digitalis toxicity (Selye et al., 1969; Yeh and Lucchesi, 1974; Waldorff and Buch, 1979). Canrenone has also been shown to reduce BP and to restore Na+/K+-ATPase activity in rats with experimental volume-dependent hypertension (de Mendonça et al., 1988; Pamnani et al., 1990). Clinical data also demonstrate that canrenone is capable of antagonizing the effects of CTS. Boero et al. (1989) have demonstrated in hypertensive subjects that canrenone can prevent inhibition of red blood cell Na+/K+-ATPase induced by intravenous saline infusion. Later, Semplicini et al. (1993) demonstrated that a 4-week administration of canrenone (100 mg daily) abolished vasoconstriction induced by a bolus infusion of ouabain in 50% of untreated hypertensive patients.
In those patients in whom ouabain-induced vasoconstriction was inhibited, canrenone significantly lowered blood pressure. In the other patients the blood pressure reduction was not statistically significant.
The next obvious approach, which also does not discriminate between the classic and “signaling” schemas of the Na+/K+-ATPase, would be the use of receptor “antagonists.” Extremely interesting data exist for PST2238 (rostafuroxin) (Fig. 3). This is a steroid compound designed to serve as an EO antagonist. Rostafuroxin has been demonstrated to have beneficial effects in a rodent ouabain infusion model (Ferrari et al., 1999, 2006; Ferrandi et al., 2004). The efficacy of rostafuroxin in hypertensive subjects and the possible dependence of its efficacy on adducin cytoskeleton proteins is being assessed in the ongoing Ouabain and Adducin for Specific Intervention on Sodium in HyperTension (OASIS-HT) (http:// clinicaltrials.gov/ct2/show/NCT00415038) phase II multicenter study (Staessen et al., 2005). Vu et al. (2006) reported that resibufagenin, a bufadienolide sodium pump inhibitor, in pregnant rats reverses hypertension induced by deoxycorticosterone acetate-salt and by chronic marinobufagenin administration.
D. Antagonism of Cardiotonic Steroid-Induced Cell Signaling
If we accept the Na+/K+-ATPase-Src-EGFR paradigm for CTS signaling, several rather obvious targets present themselves for consideration. A number of EGFR antagonists, both at an antibody (immunoneutralization) and a tyrosine kinase level, have been developed because of the importance of the EGFR for cancer. Application of these agents may be surprisingly effective in the management of cardiovascular disease. The role of reactive oxygen species in Na+/K+-ATPase signaling is another obvious target.
Neutralization of ROS may be effective in several experimental models of cardiovascular disease although the clinical benefits of this strategy are still debated.
Another possible approach to interfere with the effect of CTS is inhibition of Na+/Ca2+ exchanger, NCX1.
CTS inhibit Na+/K+-ATPase in vascular smooth muscle cells; the elevation of local Na+ facilitates Ca2+ entry through NCX1, resulting in vasoconstriction (Iwamoto, 2006). The Na+/Ca2+ exchange inhibitors having a benzyloxyphenyl structure interact with a specific receptor site in the NCX1 molecule and block its reverse (Ca2+ influx mode) mode more effectively than the forward Ca2+ efflux mode (Iwamoto et al., 2004). Thus, the benzyloxyphenyl inhibitors exhibit their effect under pathological conditions (high Na+i) rather than under normal (low Na+i) conditions. In consideration of such a profile, benzyloxyphenyl inhibitors appear promising as therapeutic agents for the treatment of conditions associated with intracellular Na+i, such as salt-dependent hypertension, in which levels of CTS are elevated. Indeed, the administration of NCX1 inhibitors, SEA0400 and KB-R7942, lowered arterial pressure in several rat models of salt-sensitive hypertension, in which levels of CTS are elevated and contribute to enhanced vascular tone in deoxycorticosterone acetate-salt hypertensive rats, salt-loaded Dahl-S rats, and rats with ACTH-induced hypertension. Remarkably, NSX1 inhibitors exhibited minimal vasodepressor effects in normal animals and in saltindependent forms of hypertension (Iwamoto et al., 2004; Iwamoto, 2006).
E. Biosynthesis and Release of Cardiotonic Steroids
One potential opportunity is to decrease the circulating concentrations of CTS. Unfortunately, we are somewhat limited in achieving this end because we do not fully understand the biosynthesis of the CTS. That said, it will be interesting to note how much of the therapeutic “accomplishments” of inhibiting the RAS are related, in fact, to the ATII dependence of the EO-MBG axis (Fig.
4). The benefits of inhibiting ATII synthesis or action have been well established in hypertension, renal failure progression, and CHF. Unfortunately, although chronic angiotensin-converting enzyme inhibition was shown to reduce levels of circulating CTS in young spontaneously hypertensive rats (Kähönen et al., 1995) and blockade of ATII receptors by losartan prevented NaCl-induced stimulation of MBG in the adrenal cortex (Fedorova et al., 2005a), we do not have robust clinical data about the long-term effects of RAS inhibition on the EO-MBG axis.
F. Conclusions and Perspectives
Our understanding of the role of endogenous cardiotonic steroids has evolved considerably. More than 50 years ago, it was hypothesized that digitalis glycosides activity and represent a potential therapeutic target.
Bagrov et al. (2000) have demonstrated that cicletanine, a furopyridine antihypertensive compound with natriuretic and vasorelaxant actions, directly inhibits PKC activity and antagonizes the vasoconstrictor and Na+/ K+-ATPase inhibitory effects of MBG via a PKC-sensitive mechanism. In a subsequent study, in Dahl-S rats with NaCl-induced hypertension, in which levels of MBG are elevated and contribute to vasospasm, chronic administration of cicletanine prevented development of hypertension, reduced the MBG sensitivity of the Na+/ K+-ATPase in LV myocardium, reduced the level of phosphorylation of the myocardial α1 Na+/K+-ATPase, and prevented up-regulation of PKC β2 and ␦-isoforms in LV sarcolemma (Fedorova et al., 2003).
2. Protein Kinase G. ANP is capable of modulating CTS sensitivity of the Na+/K+-ATPase via PKG-dependent phosphorylation. cGMP regulates activity of the Na+/K+-ATPase by modulation of its phosphorylation (Blanco et al., 1998; Fotis et al., 1999) and activation of the guanylyl cyclase-cGMP-PKG pathway underlies both renal and vascular effects of ANPs (Vesely et al., 1987; Hedge et al., 1989; Scavone et al., 1995). Unlike PKC, which directly phosphorylates the Na+/K+-ATPase, the effect of PKG on sodium pump phosphorylation is more complex and involves the recruitment and regulation of additional phosphatases (Aperia et al., 1994; Fotis et al., 1999). Fedorova et al. (2006) have shown that α-hANP and prepro-ANP, via a cGMP-dependent mechanism, dephosphorylate Na+/K+-ATPase from vascular sarcolemma and markedly reduce its sensitivity to MBG. In the same study, ANP exhibited an opposite effect; i.e., it induced Na+/K+-ATPase phosphorylation and sensitized the sodium pump to the inhibitory effect of MBG in renal medulla. Because aortic sarcolemma and renal medulla express PKG1 and PKG2 isoforms, respectively, these two PKG isoforms are likely to mediate the opposing effects of ANP on Na+/K+ATPase phosphorylation and MBG sensitivity (Fedorova et al., 2006). Thus, ANP may be capable of antagonizing undesirable effects of CTS while potentiating desired effects in some settings. Accordingly, plasma levels of MBG and α-hANP exhibit a strong positive correlation in hypertensive patients with heart failure (Fridman et al., 2002). As our understanding of this signaling pathway expands, additional targets for clinical intervention will become apparent.
sodium transport and are, rather, more analogous to that of receptor tyrosine kinases. Unfortunately, the measurement of CTS is mainly based on “in-house” immunoassays, which makes the comparison of results obtained in different laboratories difficult (Manunta et al., 2009; Nicholls et al., 2009). We propose that the development of reliable commercial methods of measurement of CTS levels is critical for the future advancement of this field.
The α-subunit of the Na+/K+-ATPase represents the specific receptor for CTS, and α isoforms of Na+/K+ATPase exhibit differential sensitivity to various CTS.
Therefore, this family of isoforms essentially can be viewed as multiple receptors for multiple endogenous CTS. Accordingly, changes in the levels of endogenous CTS are associated with levels of expression of Na+/K+ATPase as well as with the changes in its sensitivity to endogenous ligands. Although radiolabeled ouabain and digoxin are widely used in the studies of CTS, the absence of commercially available labeled bufadienolides remains a limiting factor in the studies of receptor functions of the Na+/K+-ATPase.
Although our understanding of the functions of CTS is still quite incomplete, the importance of this class of hormones seems to be considerable. Endogenous CTS exhibit physiological functions that go far beyond regulation of sodium transport, natriuresis, and blood pressure and include regulation of cell growth and differentiation, apoptosis and proliferation, and glucose metabolism and control of central nervous functions. Dysregulation of these hormones seems to play an important role in a number of disease states ranging from hypertension to cancer. We propose that expanding our understanding of this class of hormones will lead to novel and effective therapeutic strategies of great relevance to optimizing health and curing diseases.
Acknowledgments
This work was supported by the Intramural Research Program of the National Institutes of Health National Institute on Aging.
References
| # | Year | Authors | Title |
|---|---|---|---|
| 1 | Adair CD, Buckalew V, Taylor K, Ernest JM, Frye AH, Evans C, and Veille JC (1996) Elevated endoxin-like factor complicating a multifetal second trimester pregnancy: treatment with digoxin-binding immunoglobulin. Am J Nephrol 16:529 – 531. [CrossRef] [PubMed] | ||
| 2 | Adair CD, Luper A, Rose JC, Russell G, Veille JC, and Buckalew VM (2009) The hemodynamic effects of intravenous digoxin-binding Fab immunoglobulin in severe preeclampsia: a double-blind, randomized, clinical trial. J Perinatol doi: 10.1038/jp.2008.224 Adams RJ, Schwartz A, Grupp G, Grupp I, Lee SW, Wallick ET, Powell T, Twist VW, and Gathiram P (1982) High-affinity ouabain binding site and low-dose positive inotropic effect in rat myocardium. Nature 296:167–169. [CrossRef] [PubMed] | ||
| 3 | Agnoletti G, Rodella A, Ferrari R, and Harris (1987) Release of atrial natriuretic peptide-like immunoreactive material during stretching of the rat atrium in vitro. J Mol Cell Cardiol 19:217–220. [CrossRef] [PubMed] | ||
| 4 | Akera T and Brody TM (1977) The role of Na,K-ATPase in the inotropic action of digitalis. Pharmacol Rev 29:187–220. Akimova OA, Bagrov AY, Lopina OD, Kamernitsky AV, Tremblay J, Hamet P, and [CrossRef] [PubMed] | ||
| 5 | Orlov SN (2005) Cardiotonic steroids differentially affect intracellular Na+ and [Na+]i/[K+]i-independent signaling in C7-MDCK cells. J Biol Chem 280:832– 839. [CrossRef] [PubMed] | ||
| 6 | Akimova OA, Hamet P, and Orlov SN (2008a) [Na+]i/[K+]i-independent death of ouabain-treated renal epithelial cells is not mediated by Na+,K+-ATPase internalization and de novo gene expression. Pflugers Arch 455:711–719. [CrossRef] [PubMed] | ||
| 7 | 1992 | Akimova OA, Poirier M, Kotelevtsev SV, Hamet P, and Orlov SN (2008b) The death represented the exogenous counterparts of endogenous factor(s) involved in the regulation of cardiovascular system (Rein, 1949; Szent-Gyorgyi, 1953) | When it became clear that the Na+/K+-ATPase was the specific receptor for digitalis, the concept of the Third Factor emerged (de Wardener and Clarkson, 1985). Within a few years, many studies demonstrated that these endogenous CTS did exist and implicated them in the regulation of the cardiovascular system, fluid-electrolyte homeostasis, and the pathogenesis of essential hypertension (Goto et al., 1992; Schoner, 1992). The discovery of EO was a modern breakthrough in these studies of CTS (Hamlyn et al., 1991), and the discovery of the cell-signaling functions of the Na+/K+ATPase, along with the definitive chemical characterization of both cardenolide and bufadienolide CTS, has largely erased skepticism and reawakened interest in this topic. At present, it is clear that multiple endogenous Na+/K+ ATPase inhibitors exist in mammals. One of the factors underlying skepticism concerning endogenous CTS is the marked differences from their pharmacological prototype, digitalis, that endogenous CTS display. First, the classic pharmacology of endogenous CTS was based on the presumption that the effects of these compounds were mediated via inhibition of transmembrane sodium transport (Braunwald and Klocke, 1965; Akera and Brody, 1977). Data reviewed elsewhere (Xie and Askari, 2002; Schoner and ScheinerBobis, 2007; Tian and Xie, 2008) and briefly in this article, clearly demonstrate that many signaling effects of CTS may be independent of this mechanism. Second, many classic pharmacological studies of digitalis used micromolar concentrations of these agents, whereas circulating concentrations of endogenous CTS, at least in mammals, seem to be in the subnanomolar range. Very different effects of these lower concentrations of CTS on cell signaling have been observed (Nesher et al., 2007; Schoner and Scheiner-Bobis, 2007). Thus, it seems that an enormous number of studies of cardiac glycosides not only did not facilitate the progress of our understanding of endogenous CTS but also became rather an obstacle in the studies of endogenous sodium pump ligands. Despite controversies and periods of skepticism (Manunta et al., 2009; Nicholls et al., 2009), it is clear that endogenous CTS represent an important class of hormones with profound consequences in health and disease, related but not limited to their roles as natriuretic hormones. These hormones are synthesized in the adrenal cortex, and, similar to neurosteroids, in the brain. Endogenous CTS circulate bound to transport proteins, and levels of CTS in plasma vary from subnanomolar to nanomolar concentrations in different physiological conditions. These circulating levels of CTS seem to be both sufficient to alter transmembrane sodium transport in some cell types as well as to induce a variety of cellular signals, some of which seem to be independent of changes in transmembrane A (2001) Increased circulating levels of ouabain-like factor in patients with asymptomatic left ventricular dysfunction. Eur J Heart Fail 3:165–171. [CrossRef] [PubMed] |
| 8 | Balzan S, Nicolini G, Iervasi A, Di Cecco P, and Fommei E (2005) Endogenous ouabain and acute salt loading in low-renin hypertension. Am J Hypertens 18: 906 –909. Bauer N, Müller-Ehmsen J, Krämer U, Hambarchian N, Zobel C, Schwinger RH, [CrossRef] [PubMed] | ||
| 9 | Neu H, Kirch U, Grünbaum EG, and Schoner W (2005) Ouabain-like compound changes rapidly on physical exercise in humans and dogs: effects of β-blockade and angiotensin-converting enzyme inhibition. Hypertension 45:1024 –1028. [CrossRef] [PubMed] | ||
| 10 | Bergdahl B, Dahlström G, Molin L, and Bertler A (1979) Inter and intra laboratory variation of digoxin radioimmunoassay in Sweden. Acta Pharmacol Toxicol (Copenh) 45:66 –72. [CrossRef] [PubMed] | ||
| 11 | Bergdahl B and Molin L (1981) Precision of digoxin radioimmunoassays and matrix effects: four kits compared. Clin Biochem 14:67–71. [CrossRef] [PubMed] | ||
| 12 | Bernini G, Paci A, Sgrò M, Moretti A, and Salvetti A (1998) Endogenous digitalis-like factor and ouabain immunoreactivity in adrenalectomized patients and normal subjects after acute and prolonged salt loading. Am J Hypertens 11:1–7. [CrossRef] [PubMed] | ||
| 13 | Bertorello AM, Aperia A, Walaas SI, Nairn AC, and Greengard P (1991) Phosphorylation of the catalytic subunit of Na+,K+-ATPase inhibits the activity of the enzyme. Proc Natl Acad Sci U S A 88:11359 –11362. [CrossRef] [PubMed] | ||
| 14 | Bertorello AM and Katz AI (1993) Short-term regulation of renal Na-K-ATPase activity: physiological relevance and cellular mechanisms. Am J Physiol 265: F743–F755. Bianchi G, Tripodi G, Casari G, Salardi S, Barber BR, Garcia R, Leoni P, Torielli L, [CrossRef] [PubMed] | ||
| 15 | 1994 | Cusi D, Ferrandi M, et al | ( 1994 ) Two point mutations within the adducin genes are involved in blood pressure variation. Proc Natl Acad Sci U S A 26:3999 – 4003. [CrossRef] [PubMed] |
| 16 | Blanco G, Koster JC, Sánchez G, and Mercer RW (1995) Kinetic properties of the α2β1 and α2β2 isozymes of the Na,K-ATPase. Biochemistry 34:319 –325. [CrossRef] [PubMed] | ||
| 17 | Blanco G, Melton RJ, Sánchez G, and Mercer RW (1999) Functional characterization of a testes-specific α-subunit isoform of the sodium/potassium adenosinetriphosphatase. Biochemistry 38:13661–13669. [CrossRef] [PubMed] | ||
| 18 | Blanco G and Mercer RW (1998) Isozymes of the Na/K-ATPase: heterogeneity in structure, diversity in function. Am J Physiol 275:F633–F650. [CrossRef] [PubMed] | ||
| 19 | Blanco G, Sánchez G, and Mercer RW (1998) Differential regulation of Na,K-ATPase isozymes by protein kinases and arachidonic acid. Arch Biochem Biophys 359:139 – 150. [CrossRef] [PubMed] | ||
| 20 | Blaustein MP (1993) Physiological effects of endogenous ouabain: control of intracellular Ca2+ stores and cell responsiveness. Am J Physiol 264:C1367–1387. Boero R, Guarena C, Deabate MC, Rolando B, Rosati C, Quarello F, and Piccoli G (1989) Erythrocyte Na+,K+ pump inhibition after saline infusion in essentially hypertensive subjects: effects of canrenone administration. Int J Cardiol 25 (Suppl 1):S47–S52. [CrossRef] [PubMed] | ||
| 21 | Book CB, Moore RL, Semanchik A, and Ng YC (1994) Cardiac hypertrophy alters expression of Na+,K+-ATPase subunit isoforms at mRNA and protein levels in rat myocardium. J Mol Cell Cardiol 25:591– 600. [CrossRef] [PubMed] | ||
| 22 | Bourgoignie JJ, Weisser F, Rolf D, Klahr S, and Bricker NS (1970) Demonstration of a low-molecular-weight natriuretic factor in uremic serum. Trans Assoc Am Physicians 83:277–287. [CrossRef] [PubMed] | ||
| 23 | Braunwald E and Klocke FJ (1965) Digitalis. Annu Rev Med 16:371–386. [CrossRef] [PubMed] | ||
| 24 | Bricker NS, Schmidt RW, Favre H, Fine L, and Bourgoignie JJ (1975) On the biology of sodium excretion: the search for natriuretic hormone. Yale J Biol Med 48:293– 303. Briones AM, Xavier FE, Arribas SM, González MC, Rossoni LV, Alonso MJ, and [CrossRef] [PubMed] | ||
| 25 | Salaices M (2006) Alterations in structure and mechanics of resistance arteries from ouabain-induced hypertensive rats. Am J Physiol Heart Circ Physiol 291: H193–H201. [CrossRef] [PubMed] | ||
| 26 | Bruneau BG, Piazza LA, and de Bold AJ (1997) BNP gene expression is specifically modulated by stretch and ET-1 in a new model of isolated rat atria. Am J Physiol 273:H2678 –H2686. [CrossRef] [PubMed] | ||
| 27 | Buckalew VM Jr, Martinez FJ, and Green W (1970) The effect of dialysates and ultrafiltrates of plasma and saline-loaded dogs on toad bladder sodium transport. J Clin Invest 49:926 –935. [CrossRef] [PubMed] | ||
| 28 | Burns EL, Nicholas RA, and Price EM (1996) Random mutagenesis of the sheep Na,K-ATPase α1 subunit generating the ouabain-resistant mutant L793P. J Biol Chem 271:15879 –15883. Butler VP Jr, Morris JF, Akizawa T, Matsukawa M, Keating P, Hardart A, and [CrossRef] [PubMed] | ||
| 29 | Furman I (1996) Heterogeneity and lability of endogenous digitalis-like substances in the plasma of the toad, Bufo marinus. Am J Physiol 271:R325–R332. [CrossRef] [PubMed] | ||
| 30 | Butt AN, Semra YK, Ho CS, and Swaminathan R (1997) Effect of high salt intake on plasma and tissue concentration of endogenous ouabain-like substance in the rat. Life Sci 61:2367–2373. [CrossRef] [PubMed] | ||
| 31 | Cai H, Wu L, Qu W, Malhotra D, Xie Z, Shapiro JI, and Liu J (2008) Regulation of apical NHE3 trafficking by ouabain-induced activation of the basolateral Na+-K+ATPase receptor complex. Am J Physiol Cell Physiol 294:C555–C563. [CrossRef] [PubMed] | ||
| 32 | Carroll JS, Seely EW, Tao QF, and Graves SW (2001) Digitalis-like factor response to hyperinsulinemia accompanying a euglycemic hyperinsulinemic clamp or oral glucose tolerance test. Life Sci 69:829 – 837. Charlemagne D, Orlowski J, Oliviero P, Rannou F, Sainte Beuve C, Swynghedauw B, and Lane LK (1994) Alteration of Na,K-ATPase subunit mRNA and protein levels in hypertrophied rat heart. J Biol Chem 269:1541–1547. [CrossRef] [PubMed] | ||
| 33 | Chen C and Osuch MV (1969) Biosynthesis of bufadienolides-3-β-hydroxycholanates as precursors in Bufo marinus bufadienolides synthesis. Biochem Pharmacol 18: 1797–1802. [CrossRef] [PubMed] | ||
| 34 | Chen KK and Kovaríková A (1967) Pharmacology and toxicology of toad venom. J Pharm Sci 56:1535–1541. [CrossRef] [PubMed] | ||
| 35 | Chen S, Yuan C, Clough D, Schooley J, Haddy FJ, and Pamnani MB (1993) Role of digitalis-like substance in the hypertension of streptozotocin-induced diabetes in reduced renal mass rats. Am J Hypertens 6:397– 406. [CrossRef] [PubMed] | ||
| 36 | Cheung WJ, Kent MA, El-Shahat E, Wang H, Tan J, White R, and Leenen FH (2006) Central and peripheral renin-angiotensin systems in ouabain-induced hypertension. Am J Physiol Heart Circ Physiol 291:H624 –H630. of ouabain-treated renal epithelial cells: evidence against anoikis occurrence. Apoptosis 13:670 – 680. [CrossRef] [PubMed] | ||
| 37 | Albers RW (1967) Biochemical aspects of active transport. Annu Rev Biochem 36: 727–756. [CrossRef] [PubMed] | ||
| 38 | Amin MS, Wang HW, Reza E, Whitman SC, Tuana BS, and Leenen FH (2005) Distribution of epithelial sodium channels and mineralocorticoid receptors in cardiovascular regulatory centers in rat brain. Am J Physiol Regul Integr Comp Physiol 289:R1787–R1797. Amler E, Cester N, Salvolini E, Staffolani R, Burkhard M, Mazzanti L, Kotyk A, and [CrossRef] [PubMed] | ||
| 39 | Romanini C (1994) Human hypertensive placenta contains an increased amount of Na,K-ATPase with higher affinity for cardiac glycosides. Cell Biol Int 18:723–727. [CrossRef] [PubMed] | ||
| 40 | Anderson DE, Bagrov AY, and Austin JL (1995) Inhibited breathing decreases renal sodium excretion. Psychosom Med 57:373–380. Anderson DE, Fedorova OV, Morrell CH, Kashkin VA, Bagrov AY, and Lakatta EG (2008) Endogenous sodium pump inhibitors are linked to age-associated increases in salt sensitivity of blood pressure in normotensives. Am J Physiol Regul Integr Physiol 294:R1248 –R1254. Antolovic R, Bauer N, Mohadjerani M, Kost H, Neu H, Kirch U, Grünbaum EG, and [CrossRef] [PubMed] | ||
| 41 | Schoner W (2000) Endogenous ouabain and its binding globulin: effects of physical exercise and study on the globulin’s tissue distribution. Hypertens Res 23 (Suppl): S93–S98. [CrossRef] [PubMed] | ||
| 42 | Antolovic R, Kost H, Mohadjerani M, Linder D, Linder M, and Schoner W (1998) A specific binding protein for cardiac glycosides exists in bovine serum. J Biol Chem 273:16259 –16264. Aperia A, Holtback U, Syren ML, Svensson LB, Fryckstedt J, and Greengard P (1994) Activation/deactivation of renal Na,K-ATPase: a final common pathway for regulation of natriuresis. FASEB J 8:436 – 469. Appel LJ, Brands MW, Daniels SR, Karanja N, Elmer PJ, Sacks FM, and American [CrossRef] [PubMed] | ||
| 43 | Heart Association (2006) Dietary approaches to prevent and treat hypertension: a scientific statement from the American Heart Association. Hypertension 47:296 – 308. Averina IV, Tapilskaya NI, Reznik VA, Frolova EV, Fedorova OV, Lakatta EG, and [CrossRef] [PubMed] | ||
| 44 | Bagrov AY (2006) Endogenous Na/K-ATPase inhibitors in patients with preeclampsia. Cell Mol Biol 52:19 –23. Bagrov AY, Bagrov YY, Fedorova OV, Kashkin VA, Patkina NA, and Zvartau EE (2002) Endogenous digitalis-like ligands of the sodium pump: possible involvement in mood control and ethanol addiction. Eur Neuropsychopharmacol 12:1–12. Bagrov AY, Dmitrieva RI, Dorofeeva NA, Fedorova OV, Lopatin DA, Lakatta EG, and Droy-Lefaix MT (2000) Cicletanine reverses vasoconstriction induced by a putative endogenous Na/K ATPase ligand, marinobufagenin, via protein kinase C dependent mechanism. J Hypertens 18:209 –215. Bagrov AY, Dmitrieva RI, Fedorova OV, Kazakov GP, Roukoyatkina NI, and Shpen VM (1996a) Endogenous marinobufagenin-like immunoreactive substance: a possible endogenous Na,K-ATPase inhibitor with vasoconstrictor activity. Am J Hypertens 9:982–990. [CrossRef] [PubMed] | ||
| 45 | Bagrov AY and Fedorova OV (1998) Effects of two putative endogenous digitalis-like factors, marinobufagenin and ouabain, on the Na+,K+-pump in human mesenteric arteries. J Hypertens 16:1953–1958. [CrossRef] [PubMed] | ||
| 46 | Bagrov AY, Fedorova OV, Austin JL, Dmitrieva RI, and Anderson DE (1995a) Endogenous marinobufagenin-like immunoreactive factor and Na,K-ATPase inhibition during voluntary hypoventilation. Hypertension 26:781–788. [CrossRef] [PubMed] | ||
| 47 | Bagrov AY, Fedorova OV, Dmitrieva RI, French AW, and Anderson DE (1996b) Plasma marinobufagenin-like and ouabain-like immunoreactivity during acute saline volume expansion in anesthetized dogs. Cardiovasc Res 206:296 –305. Bagrov AY, Fedorova OV, Dmitrieva RI, Howald WN, Hunter AP, Kuznetsova EA, and Shpen VM (1998) Characterization of a urinary bufodienolide Na+,K+-ATPase inhibitor in patients after acute myocardial infarction. Hypertension 31:1097– 1103. Bagrov AY, Fedorova OV, Maslova MN, Roukoyatkina NI, Stolba P, and Zhabko EP (1989) Antiarrhythmic effect of antibodies to digoxin in acute myocardial ischemia in rats. Eur J Pharmacol 162:195–196. Bagrov AY, Fedorova OV, Maslova MN, Roukoyatkina NI, Ukhanova MV, and [CrossRef] [PubMed] | ||
| 48 | Zhabko EP (1991) Endogenous plasma Na,K-ATPase inhibitory activity and digoxin-like immunoreactivity after acute myocardial infarction. Cardiovasc Res 25: 371–377. [CrossRef] [PubMed] | ||
| 49 | Bagrov AY, Fedorova OV, Roukoyatkina NI, Ukhanova MV, and Zhabko EP (1993a) Digitalis-like and vasoconstrictor properties of endogenous digoxin-like factor from Bufo marinus toad. Eur J Pharmacol 234:165–172. [CrossRef] [PubMed] | ||
| 50 | Bagrov AY, Fedorova OV, Roukoyatkina NI, and Zhabko EP (1993b) Effect of antidigoxin antibody on myocardial Na,K-pump activity and of endogenous digoxin-like factor in acute myocardial ischemia in rats. Cardiovasc Res 27:1045–1050. [CrossRef] [PubMed] | ||
| 51 | Bagrov AY, Kuznetsova EA, Fedorova OV (1994) Endogenous digoxin-like factor in acute myocardial infarction. J Intern Med 235:63– 67. [CrossRef] [PubMed] | ||
| 52 | Bagrov AY, Roukoyatkina NI, Dmitrieva RI, Pinaev AG, and Fedorova OV (1995b) Effects of two endogenous digitalis-like factors, ouabain and marinobufagenin in isolated rat aorta. Eur J Pharmacol 274:151–158. [CrossRef] [PubMed] | ||
| 53 | Bagrov YY, Dmitrieva NI, and Manusova NB (1996c) Competition of ethanol with endogenous digoxin-like factor as a possible mechanism of addiction. Dokl Akad Nauk (Proc Russ Acad Sci) 348: 268 –269 (in Russian). Bagrov YY, Dmitrieva NI, Manusova NB, Zvartau EE, Patkina NA and Bagrov AY (1999) Possible role of endogenous digitalis-like factor in the voluntary selection of alcohol by rats. Life Sci 64: PL219 –PL225. [CrossRef] [PubMed] | ||
| 54 | Bagrov YY, Manusova NB, Egorova IA, Fedorova OV, and Bagrov AY (2005) Endogenous digitalis-like substances and Na/K-ATPase inhibition in diabetes mellitus. Front Biosci 10:2257–2262. Bagrov YY, Manusova NB, Frolova EV, Egorova IA, Kashkin VA, Tapilskaya NI, [CrossRef] [PubMed] | ||
| 55 | Fedorova OV, and Bagrov AY (2007) Endogenous sodium pump inhibitors, diabetes mellitus and preeclampsia: preliminary observations and a hypothesis. Pathophysiology 14:147–151. Balzan S, Neglia D, Ghione S, D’Urso G, Baldacchino MC, Montali U, and L’Abbate + Na ,K -ATPase trafficking in response to GPCR signals and intracellular sodium. Circ Res 95:1100 –1108. [CrossRef] [PubMed] | ||
| 56 | Ehlenz K, Peter JH, Kaffarnik H, and von Wichert P (1991) Disturbances in volume regulating hormone system—a key to the pathogenesis of hypertension in obstructive sleep apnea syndrome? Pneumologie 45 (Suppl 1):239 –245. Elkareh J, Kennedy DJ, Yashaswi B, Vetteth S, Shidyak A, Kim EG, Smaili S, [CrossRef] [PubMed] | ||
| 57 | 2007 | Periyasamy SM, Hariri IM, Fedorova L, et al | ( 2007 ) Marinobufagenin stimulates fibroblast collagen production and causes fibrosis in experimental uremic cardiomyopathy. Hypertension 49:215–224. el-Mallakh RS, Harrison LT, Li R, Changaris DG, and Levy RS (1995) An animal model for mania: preliminary results. Progr Neuropsychopharmacol Biol Psychiatry 19:955–962. el-Mallakh RS and Wyatt RJ (1995) The Na,K-ATPase hypothesis for bipolar illness. Biol Psychiatry 37:235–244. el-Masri MA, Clark BJ, Qazzaz HM, and Valdes R Jr (2002) Human adrenal cells in culture produce both ouabain-like and dihydroouabain-like factors. Clin Chem 48:1720 –1730. [CrossRef] [PubMed] |
| 58 | 2008 | El-Okdi N, Smaili S, Raju V, Shidyak A, Gupta S, Fedorova L, Elkareh J, Periyasamy S, Shapiro AP, Kahaleh MB, et al | ( 2008 ) Effects of cardiotonic steroids on dermal collagen synthesis and wound healing. J Appl Physiol 105:30 –36. [CrossRef] [PubMed] |
| 59 | Fambrough DM (1988) The sodium pump becomes a family. Trends Neurosci 11: 325–328. [CrossRef] [PubMed] | ||
| 60 | Farley RA, Heart E, Kabalin M, Putnam D, Wang K, Kasho VN, and Faller LD (1997) Site-directed mutagenesis of the sodium pump: analysis of mutations to amino acids in the proposed nucleotide binding site by stable oxygen isotope exchange. Biochemistry 36:941–951. [CrossRef] [PubMed] | ||
| 61 | Fedorova OV, Agalakova NI, Morrell CH, Lakatta EG, and Bagrov AY (2006) ANP differentially modulates marinobufagenin-induced sodium pump inhibition in kidney and aorta. Hypertension 48:1160 –1168. [CrossRef] [PubMed] | ||
| 62 | Fedorova OV, Agalakova NI, Talan MI, Lakatta EG, and Bagrov AY (2005a) Brain ouabain stimulates peripheral marinobufagenin via angiotensin II signalling in NaCl loaded Dahl-S rats. J Hypertens 23:1515–1523. [CrossRef] [PubMed] | ||
| 63 | Fedorova OV, Anderson DE, and Bagrov AY (1998) Endogenous digitalis-like factors and Na,K-ATPase inhibition in ACTH hypertension in rats. Am J Hypertens 11:796 – 802. [CrossRef] [PubMed] | ||
| 64 | Fedorova OV, Anderson DE, Lakatta EG, and Bagrov AY (2001a) Interaction of high sodium chloride intake and psychosocial stress on endogenous ligands of the sodium pump and blood pressure in normotensive rats. Am J Physiol 281:R352– R358. [CrossRef] [PubMed] | ||
| 65 | Fedorova OV and Bagrov AY (1997) Inhibition of Na/K ATPase from rat aorta by two endogenous Na/K pump inhibitors, ouabain and marinobufagenin. Evidence of interaction with different alpha-subinit isoforms. Am J Hypertens 10:929 –935. [CrossRef] [PubMed] | ||
| 66 | Fedorova OV, Doris PA, and Bagrov AY (1998) Endogenous marinobufagenin-like factor in acute plasma volume expansion. Clin Exp Hypertens 20:581–591. [CrossRef] [PubMed] | ||
| 67 | Fedorova OV, Dorofeeva NA, Lopatin DA, Lakatta EG, and Bagrov AY (2002a) Phorbol diacetate potentiates Na/K-ATPase inhibition by a putative endogenous ligand, marinobufagenin. Hypertension 39:298 –302. [CrossRef] [PubMed] | ||
| 68 | Fedorova OV, French AW, and Anderson DE (1996) Inhibition of erythrocyte Na,KATPase activity during anticipatory hypoventilation in micropigs. Am J Hypertens 9:1126 –1131. [CrossRef] [PubMed] | ||
| 69 | Fedorova OV, Kolodkin NI, Agalakova NI, Lakatta EG, and Bagrov AY (2001b) Marinobufagenin, an endogenous α-1 sodium pump ligand, in hypertensive Dahl salt-sensitive rats. Hypertension 37:462– 466. Fedorova OV, Kolodkin NI, Agalakova NI, Namikas AR, Bzhelyansky A, St-Louis J, [CrossRef] [PubMed] | ||
| 70 | Lakatta EG, and Bagrov AY (2005b) Antibody to marinobufagenin lowers blood pressure in pregnant rats on a high NaCl intake. J Hypertens 23:835– 842. [CrossRef] [PubMed] | ||
| 71 | Fedorova OV, Lakatta EG, and Bagrov AY (2000) Differential effects of acute NaCl loading on endogenous ouabain-like and marinobufagenin-like ligands of the sodium pump in Dahl hypertensive rats. Circulation 102:3009 –3014. Fedorova OV, Simbirtsev AS, Kolodkin NI, Kotov AY, Agalakova NI, Kashkin VA, [CrossRef] [PubMed] | ||
| 72 | 2008 | Tapilskaya NI, Bzhelyansky AM, Reznik VA, Nikitina ER, et al | ( 2008 ) Monoclonal antibody to an endogenous bufadienolide, marinobufagenin, reverses preeclampsia-induced Na/K-ATPase inhibition in lowers blood pressure in NaCl-sensitive hypertension. J Hypertens 26:2414 –2425. Fedorova OV, Talan MI, Agalakova NI, Droy-Lefaix M-T, Lakatta EG, and Bagrov AY (2003) Reduction in myocardial PKC β-2, Na/K-ATPase sensitivity to marinobufagenin and blood pressure in response to cicletanine. Hypertension 41:505– 511. [CrossRef] [PubMed] |
| 73 | Fedorova OV, Talan MI, Agalakova NI, Lakatta EG, and Bagrov AY (2002b) An endogenous ligand of α-1 sodium pump, marinobufagenin, is a novel mediator of sodium chloride dependent hypertension. Circulation 105:1122–1127. [CrossRef] [PubMed] | ||
| 74 | Fedorova OV, Talan MI, Agalakova NI, Lakatta EG, and Bagrov AY (2004) Coordinated shifts in Na/K-ATPase isoforms and their endogenous ligands during cardiac hypertrophy and failure in NaCl-sensitive hypertension. J Hypertens 22:389 –397. Fedorova OV, Zhuravin IA, Agalakova NI, Yamova LA, Talan MI, Lakatta EG, and [CrossRef] [PubMed] | ||
| 75 | Bagrov AY (2007) Intrahippocampal microinjection of an exquisitely low dose of ouabain mimics NaCl loading and stimulates a bufadienolide Na/K-ATPase inhibitor. J Hypertens 25:1834 –1844. Feldmann T, Glukmann V, Medvenev E, Shpolansky U, Galili D, Lichtstein D, and [CrossRef] [PubMed] | ||
| 76 | Rosen H (2007) Role of endosomal Na+-K+-ATPase and cardiac steroids in the regulation of endocytosis. Am J Physiol Cell Physiol 293:C885–C896. Ferrandi M, Barassi P, Molinari I, Torielli L, Tripodi G, Minotti E, Bianchi G, and [CrossRef] [PubMed] | ||
| 77 | Ferrari P (2005) Ouabain antagonists as antihypertensive agents. Curr Pharm Des 11:3301–3305. [CrossRef] [PubMed] | ||
| 78 | Ferrandi M, Minotti E, Salardi S, Florio M, Bianchi G, and Ferrari P (1993) Characteristics of a ouabain-like factor from Milan hypertensive rats. J Cardiovasc Pharmacol 22 (Suppl 2):S75–S78. [CrossRef] [PubMed] | ||
| 79 | Ferrandi M, Molinari I, Barassi P, Minotti E, Bianchi G, and Ferrari P (2004) Organ hypertrophic signaling within caveolae membrane subdomains triggered by ouabain and antagonized by PST 2238. J Biol Chem 279:33306 –33314. Ferrari P, Ferrandi M, Tripodi G, Torielli L, Padoani G, Minotti E, Melloni P, and Chibalin AV, Ogimoto G, Pedemonte CH, Pressley TA, Katz AI, Féraille E, Berggren [CrossRef] [PubMed] | ||
| 80 | PO, and Bertorello AM (1999) Dopamine-induced endocytosis of Na+,K+-ATPase is initiated by phosphorylation of Ser-18 in the rat α subunit and is responsible for the decreased activity in epithelial cells. J Biol Chem 274:1920 –1927. [CrossRef] [PubMed] | ||
| 81 | Choi SJ, Taylor MA, and Abrams R (1977) Depression, ETC and erythrocyte adenosine triphosphatase activity. Biol Psychiatry 12:75– 81. [CrossRef] [PubMed] | ||
| 82 | Clerico A, Cambi A, Del Chicca MG, Cecchini L, and Giaconi S (1988) Urinary excretion of digoxin-like immunoreactivity after physical exercise. Clin Chem 34:215. [CrossRef] [PubMed] | ||
| 83 | Clerico A and Giampietro O (1990) Is the endogenous digitalis-like factor the link between hypertension and metabolic disorders as diabetes mellitus, obesity and acromegaly? Clin Physiol Biochem 8:153–168. [CrossRef] [PubMed] | ||
| 84 | Cornelius F and Mahmmoud YA (2003) Direct activation of gastric H,K-ATPase by N-terminal protein kinase C phosphorylation: comparison of the acute regulation mechanisms of H,K-ATPase and Na,K-ATPase. Biophys J 84:1690 –1700. [CrossRef] [PubMed] | ||
| 85 | Cort JH and Lichardus B (1963) The natriuretic activity of jugular vein blood during carotid occlusion. Physiol Bohemoslov 12:497–501. [CrossRef] [PubMed] | ||
| 86 | Coughlin SR, Mawdsley L, Mugarza JA, Calverley PM, and Wilding JP (2004) Obstructive sleep apnoea is independently associated with an increased prevalence of metabolic syndrome. Eur Heart J 25:735–741. Crambert G, Balzan S, Paci A, Decollogne S, Montali U, Ghione S, and Lelièvre LG (1998) Inhibition of rat Na+/K+-ATPase isoforms by endogenous digitalis extracts from neonatal human plasma. Clin Exp Hypertens 20:669 – 674. [CrossRef] [PubMed] | ||
| 87 | Crambert G and Geering K (2003) FXYD proteins: new tissue-specific regulators of the ubiquitous Na,K-ATPase. Sci STKE 2003: RE1. [CrossRef] [PubMed] | ||
| 88 | Croyle ML, Woo AL, and Lingrel JB (1997) Extensive random mutagenesis analysis of the Na+/K+-ATPase alpha subunit identifies known and previously unidentified amino acid residues that alter ouabain sensitivity-implications for ouabain binding. Eur J Biochem 248:488 – 495. [CrossRef] [PubMed] | ||
| 89 | Dahl LK, Knudsen KD, and Iwai J (1969) Humoral transmission of hypertension: evidence from parabiosis. Circ Res 24 (Suppl):21–33. de Bold AJ, Borenstein HB, Veress AT, and Sonnenberg H (1981) A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Life Sci 28:89 –94. Delva P, Barbieri E, Devynck M-A, Meyer P, Destro G, Capra C, Pernollet MG, [CrossRef] [PubMed] | ||
| 90 | Zardini P, and Lechi A (1990) Increase in plasma digitalis like activity during percutaneous transluminal angioplasty in patients with coronary stenosis. Life Sci 47:385–389. de Mendonça M, Grichois ML, Pernollet MG, Wauquier I, Trouillet-Thormann B, [CrossRef] [PubMed] | ||
| 91 | Meyer P, Devynck MA, and Garay R (1988) Antihypertensive effect of canrenone in a model where endogenous ouabain-like factors are present. J Cardiovasc Pharmacol 11:75– 83. de Wardener HE and Clarkson EM (1985) Concept of natriuretic hormone. Physiol Rev 65:658 –759. de Wardener HE, Clarkson EM, Nutbourne DM, Schrier RW, Talner LB, Ventom [CrossRef] [PubMed] | ||
| 92 | MG, and Verroust PJ (1971) Evidence for a hormone other than aldosterone which controls urinary sodium excretion. Adv Nephrol Necker Hosp 1:97–111. de Wardener HE, Fabian M, Jones JJ, Lee J, Schrier RW, and Verroust PJ (1968) The effect of acute extracellular fluid volume expansion and acute hemorrhage on plasma antidiuretic hormone and oxytocin levels in dogs. J Physiol 196: 121P– 122P. [CrossRef] [PubMed] | ||
| 93 | De Wardener HE and MacGregor GA (2002) Sodium and blood pressure. Curr Opin Cardiol 17:360 –367. [CrossRef] [PubMed] | ||
| 94 | De Wardener HE, Mills IH, Clapham WF, and Hayter CJ (1961) Studies on the efferent mechanism of the sodium diuresis which follows the administration of intravenous saline in the dog. Clin Sci 21:249 –258. [CrossRef] [PubMed] | ||
| 95 | Di Grande A, Boura AL, Read MA, Malatino LS, and Walters WA (1993) Release of a substance from the human placenta having digoxin-like immunoreactivity. Clin Exp Pharmacol Physiol 20:603– 607. [CrossRef] [PubMed] | ||
| 96 | Dmitrieva RI, Bagrov AY, Lalli E, Sassone-Corsi P, Stocco DM, and Doris PA (2000) Mammalian bufadienolide is synthesized from cholesterol in the adrenal cortex by a pathway that is independent of cholesterol side-chain cleavage. Hypertension 36:442– 448. [CrossRef] [PubMed] | ||
| 97 | Dmitrieva RI, Lalli E, and Doris PA (2005) Regulation of adrenocortical cardiotonic steroid production by dopamine and PKA signaling. Front Biosci 10:2489 –2495. [CrossRef] [PubMed] | ||
| 98 | Doris PA, Jenkins LA, and Stocco DM (1994) Is ouabain an authentic endogenous mammalian substance derived from the adrenal? Hypertension 23:632– 638. [CrossRef] [PubMed] | ||
| 99 | Doris PA, Kilgore MW, Durham D, Alberts D, and Stocco DM (1989) An endogenous digitalis-factor derived from the adrenal gland: studies of adrenocortical tumor cells. Endocrinology 125:2580 –2586. Dostanic I, Lorenz JN, Schultz Jel J, Grupp IL, Neumann JC, Wani MA, and Lingrel JB (2003) The α2 isoform of Na,K-ATPase mediates ouabain-induced cardiac inotropy in mice. J Biol Chem 278:53026 –53034. [CrossRef] [PubMed] | ||
| 100 | Dostanic I, Paul RJ, Lorenz JN, Theriault S, Van Huysse JW, and Lingrel JB (2005) The α2-isoform of Na-K-ATPase mediates ouabain-induced hypertension in mice and increased vascular contractility in vitro. Am J Physiol Heart Circ Physiol 288:H477–H485. [CrossRef] [PubMed] | ||
| 101 | Dostanic I, Schultz Jel J, Lorenz JN, and Lingrel JB (2004) The α1 isoform of Na,K-ATPase regulates cardiac contractility and functionally interacts and colocalizes with the Na/Ca exchanger in heart. J Biol Chem 279:54053–54061. Dostanic-Larson I, Lorenz JN, Van Huysse JW, Neumann JC, Moseley AE, and [CrossRef] [PubMed] | ||
| 102 | Lingrel JB (2006) Physiological role of the α1and α2-isoforms of the Na+-K+ATPase and biological significance of their cardiac glycoside binding site. Am J Physiol Regul Integr Comp Physiol 290:R524 –R528. [CrossRef] [PubMed] | ||
| 103 | Dostanic-Larson I, Van Huysse JW, Lorenz JN, and Lingrel JB (2005) The highly conserved cardiac glycoside binding site of Na,K-ATPase plays a role in blood pressure regulation. Proc Natl Acad Sci U S A 102:15845–15850. Efendiev R, Krmar RT, Ogimoto G, Zwiller J, Tripodi G, Katz AI, Bianchi G, [CrossRef] [PubMed] | ||
| 104 | Pedemonte CH, and Bertorello AM (2004) Hypertension-linked mutation in the adducin alpha-subunit leads to higher AP2-mu2 phosphorylation and impaired + [CrossRef] [PubMed] | ||
| 105 | Goto A, Yamada K, Yagi N, Hui C, and Sugimoto T (1991b) Digoxin-like immunoreactivity: is it still worth measuring? Life Sci 49:1667–1678. [CrossRef] [PubMed] | ||
| 106 | Goto A, Yamada K, Yagi N, Yoshioka M, and Sugimoto T (1992) Physiology and pharmacology of endogenous digitalis-like factors. Pharmacol Rev 44:377–399. Gottlieb SS, Rogowski AC, Weinberg M, Krichten CM, Hamilton BP, and Hamlyn JM (1992) Elevated concentrations of endogenous ouabain in patients with congestive heart failure. Circulation 86:420 – 425. [CrossRef] [PubMed] | ||
| 107 | Grider G, el-Mallakh RS, Huff MO, Buss TJ, Miller J, and Valdes R Jr (1999) Endogenous digoxin-like immunoreactive factor (DLIF) serum concentrations are decreased in manic bipolar patients compared to normal controls. J Affect Disord 54:261–267. [CrossRef] [PubMed] | ||
| 108 | Graves SW (1987) The possible role of digitalis-like factors in pregnancy-induced hypertension. Hypertension 10:I84 –I86. [CrossRef] [PubMed] | ||
| 109 | Graves SW, Markides KE, and Hollenberg NK (2000) Application of supercritical fluid chromatography to characterize a labile digitalis-like factor. Hypertension 36:1059 –1064. [CrossRef] [PubMed] | ||
| 110 | Graves SW, Valdes R Jr, Brown BA, Knight AB, and Craig HR (1984) Endogenous digoxin-immunoreactive substance in human pregnancies. J Clin Endocrinol Metab 58:748 –751. Gruber KA, Whitaker JM, and Buckalew VM Jr Endogenous digitalis-like substance in plasma of volume-expanded dogs. Nature 287:743–745, 1980. [CrossRef] [PubMed] | ||
| 111 | Gusdon JP Jr, Buckalew VM Jr, and Hennessy JF (1984) A digoxin-like immunoreactive substance in preeclampsia. Am J Obstet Gynecol 150:83– 85. [CrossRef] [PubMed] | ||
| 112 | Guyton AC, Coleman TG, Young DB, Lohmeier TE, and DeClue JW (1980) Salt balance and long-term blood pressure control. Annu Rev Med 31:15–27. [CrossRef] [PubMed] | ||
| 113 | Haas M, Askari A, and Xie Z (2000) Involvement of Src and epidermal growth factor receptor in the signal-transducing function of Na+/K+-ATPase. J Biol Chem 275: 27832–27837. [CrossRef] [PubMed] | ||
| 114 | Haas M, Wang H, Tian J, and Xie Z (2002) Src-mediated inter-receptor cross-talk between the Na+/K+-ATPase and the epidermal growth factor receptor relays the signal from ouabain to mitogen-activated protein kinases. J Biol Chem 277: 18694 –18702. [CrossRef] [PubMed] | ||
| 115 | Haddy FJ (2006) Role of dietary salt in hypertension. Life Sci 79:1585–1592. [CrossRef] [PubMed] | ||
| 116 | Haddy FJ and Overbeck HW (1976) The role of humoral agents in volume expanded hypertension. Life Sci 19:935–947. [CrossRef] [PubMed] | ||
| 117 | Hamlyn JM (2004) Biosynthesis of endogenous cardiac glycosides by mammalian adrenocortical cells: three steps forward. Clin Chem 50:469 – 470. Hamlyn JM, Blaustein MP, Bova S, DuCharme DW, Harris DW, Mandel F, Mathews [CrossRef] [PubMed] | ||
| 118 | WR, and Ludens JH (1991) Identification and characterization of a ouabain-like compound from human plasma. Proc Natl Acad Sci U S A 88:6259 – 6263. Hamlyn JM, Ringel R, Schaeffer J, Levinson PD, Hamilton BP, Kowarski AA, and [CrossRef] [PubMed] | ||
| 119 | Blaustein MP (1982) A circulating inhibitor of (Na++K+)ATPase associated with essential hypertension. Nature 300:650 – 652. [CrossRef] [PubMed] | ||
| 120 | Hansen O (2003) No evidence for a role in signal-transduction of Na+/K+-ATPase interaction with putative endogenous ouabain. Eur J Biochem 270:1916 –1919. [CrossRef] [PubMed] | ||
| 121 | Harnett JD and Parfrey PS (1994) Cardiac disease in uremia. Semin Nephrol 14:245–252. [CrossRef] [PubMed] | ||
| 122 | Hauptman PJ and Kelly RA (1999) Digitalis. Circulation 99:1265–1270. [CrossRef] [PubMed] | ||
| 123 | Hedge SS, DeFeo ML, and Jandhyala BS (1989) Effects of atrial natriuretic factor on cyclic GMP content in the rat aortic smooth muscle: studies on the role of membrane Na+,K+-ATPase. Clin Exp Pharmacol Physiol 16:623– 629. [CrossRef] [PubMed] | ||
| 124 | Hesketh JE, Glen AI, and Reading HW (1977) Membrane ATPase activities in depressive illness. J Neurochem 28:1401–1402. [CrossRef] [PubMed] | ||
| 125 | Hieber V, Siegel GJ, Fink DJ, Beaty MW, and Mata M (1991) Differential distribution of (Na,K)-ATPase α isoforms in the central nervous system. Cell Mol Neurobiol 11:253–262. Hilton PJ, White RW, Lord GA, Garner GV, Gordon DB, Hilton MJ, Forni LG, [CrossRef] [PubMed] | ||
| 126 | 1996 | McKinnon W, Ismail FM, Keenan M, et al | ( 1996 ) An inhibitor of the sodium pump obtained from human placenta. Lancet 348:303–305. [CrossRef] [PubMed] |
| 127 | Hillyard SD, Lu E, and Gonick HC (1976) Further characterization of the natriuretic factor derived from kidney tissue of volume-expanded rats: effects on short-circuit current and sodium-potassium-adenosine triphosphatase activity. Circ Res 38: 250 –255. [CrossRef] [PubMed] | ||
| 128 | Ho CS, Butt A, Semra YK, and Swaminathan R (1997) Effect of carbidopa on the excretion of sodium, dopamine, and ouabain-like substance in the rat. Hypertension 30:1544 –1548. Hopate M, Graves S, Adair D, Lam G, Johnson D, Saade G, Lewis D, Porter K, [CrossRef] [PubMed] | ||
| 129 | Humphrey R, and Chauhan N (2008) In-vivo reversal of functional sodium pump inhibition with Digibind treatment, in Proceedings of the XVIth Congress of the International Society for the Study of Hypertension in Pregnancy; 2008 Sept 20 –24; Washington DC. p 69, International Society for the Study of Hypertension in Pregnancy, Rotterdam, The Netherlands. [CrossRef] [PubMed] | ||
| 130 | Howard R and Foerstl H (1990) Toad-lickers psychosis—a warning. Br J Psychiatry 157:779 –780. [CrossRef] [PubMed] | ||
| 131 | Huang BS, Amin MS, and Leenen FH (2006) The central role of the brain in salt-sensitive hypertension. Curr Opin Cardiol 21:295–304. [CrossRef] [PubMed] | ||
| 132 | Huang BS, Kudlac M, Kumarathasan R, and Leenen FH (1999) Digoxin prevents ouabain and high salt intake-induced hypertension in rats with sinoaortic denervation. Hypertension 34:733–738. [CrossRef] [PubMed] | ||
| 133 | Huang BS and Leenen FH (1996a) Brain “ouabain” and angiotensin II in saltsensitive hypertension in spontaneously hypertensive rats. Hypertension 28:1005– 1012. [CrossRef] [PubMed] | ||
| 134 | Huang BS and Leenen FH (1996b) Sympathoexcitatory and pressor responses to increased brain sodium and ouabain are mediated via brain ANGII. Am J Physiol 270:H275–H280. [CrossRef] [PubMed] | ||
| 135 | Huang BS and Leenen FH (1998) Both brain angiotensin II and “ouabain” contribute to sympathoexcitation and hypertension in Dahl S rats on high salt intake. Hypertension 32:1028 –1033. [CrossRef] [PubMed] | ||
| 136 | Huang BS, Van Vliet BN, and Leenen FH (2004) Increases in CSF [Na+] precede the increases in blood pressure in Dahl S rats and SHR on a high-salt diet. Am J Physiol Heart Circ Physiol 287:H1160 –H1166. [CrossRef] [PubMed] | ||
| 137 | Bianchi G (1999) PST 2238: a new antihypertensive compound that modulates Na,K-ATPase in genetic hypertension. J Pharmacol Exp Ther 288:1074 –1083. [CrossRef] [PubMed] | ||
| 138 | Ferrari P, Ferrandi M, Valentini G, and Bianchi G (2006) Rostafuroxin: an ouabain antagonist that corrects renal and vascular Na+-K+-ATPase alterations in ouabain and adducin-dependent hypertension. Am J Physiol Regul Integr Comp Physiol 290:R529 –R535. [CrossRef] [PubMed] | ||
| 139 | Feschenko MS and Sweadner KJ (1997) Phosphorylation of Na,K ATPase by protein kinase C at Ser18 occurs in intact cells but does not result in direct inhibition of ATP hydrolysis. J Biol Chem 272:17726 –17733. [CrossRef] [PubMed] | ||
| 140 | Feschenko MS, Wetzel RK, and Sweadner KJ (1997) Phosphorylation of Na,KATPase by protein kinases: sites, susceptibility, and consequences. Ann N Y Acad Sci 834:479 – 488. [CrossRef] [PubMed] | ||
| 141 | Finotti P and Palatini P (1981) Canrenone as a partial agonist at the digitalis receptor site of sodium-potassium-activated adenosine triphosphatase. J Pharmacol Exp Ther 217:784 –790. [CrossRef] [PubMed] | ||
| 142 | Flier J, Edwards MW, Daly JW, and Myers CW (1980) Widespread occurrence in frogs and toads of skin compounds interacting with the ouabain site of Na+,K+ATPase. Science 208:503–505. [CrossRef] [PubMed] | ||
| 143 | Foley TD and Linnoila M (1995) Nanomolar concentrations of ouabain block ethanolinducible Na,K-ATPase activity in brain. Eur J Pharmacol 292:287–292. [CrossRef] [PubMed] | ||
| 144 | Foley TD and Rhoads DE (1994) Stimulation of Na,K-ATPase by ethanol: possible involvement of an isozyme-specific inhibitor of Na/K-ATPase. Brain Res 653:167– 172. [CrossRef] [PubMed] | ||
| 145 | Foster DM, Huber MD, and Klemm WR (1989) Ethanol may stimulate or inhibit (Na++K+)-ATPase, depending upon Na and K concentrations. Alcohol 6:437– 443. [CrossRef] [PubMed] | ||
| 146 | Fotis H, Tatjanenko LV, and Vasilets LA (1999) Phosphorylation of the α-subunits of the Na+/K+-ATPase from mammalian kidneys and Xenopus oocytes by cGMPdependent protein kinase results in stimulation of ATPase activity. Eur J Biochem 260:904 –910. [CrossRef] [PubMed] | ||
| 147 | Franzin CM, Yu J, Thai K, Choi J, and Marassi FM (2005) Correlation of gene and protein structures in the FXYD family proteins. J Mol Biol 354:743–750. Fridman AI, Matveev SA, Agalakova NI, Fedorova OV, Lakatta EG, and Bagrov AY (2002) Marinobufagenin, an endogenous ligand of α-1 Na/K-ATPase, is a marker of congestive heart failure severity. J Hypertens 20:1189 –1194. [CrossRef] [PubMed] | ||
| 148 | Funai EF, Friedlander Y, Paltiel O, Tiram E, Xue X, Deutsch L, and Harlap S (2005) Long-term mortality after preeclampsia. Epidemiology 16:206 –215. Fuse K, Satoh M, Yokota T, Ohdaira T, Muramatsu Y, Suzuki E, and Arakawa M (1999) Regulation of ventilation before and after sleep in patients with obstructive sleep apnoea. Respirology 4:125–130. [CrossRef] [PubMed] | ||
| 149 | Gabor A and Leenen FH (2009) Mechanisms in the PVN mediating local and central sodium-induced hypertension in Wistar rats. Am J Physiol Regul Integr Comp Physiol doi: 10.1152/ajpregu.90417.2008. [CrossRef] [PubMed] | ||
| 150 | Gallery ED, Hunyor SM, and Gyory AZ (1979) Plasma volume contraction: a significant factor in both pregnancy-associated hypertension (pre-eclampsia) and chronic hypertension in pregnancy. Q J Med 192:593– 602. [CrossRef] [PubMed] | ||
| 151 | Garraffo HM and Gros EG (1986) Biosynthesis of bufadienolides in toads. VI. Experiments with [1,2–3H]cholesterol, [2114 C]coprostanol, and 5 β -[2114 C]pregnenolone in the toad Bufo arenarum. Steroids 48:251–257. [CrossRef] [PubMed] | ||
| 152 | Garay RP, Diez J, Nazaret C, Dagher G, and Abitbol JP (1985) The interaction of canrenone with the Na+,K+ pump in human red blood cells. Naunyn Schmiedebergs Arch Pharmacol 329:311–315. Gault MH, Vasdev S, Longerich L, Johnson E, Farid N, Legal Y, Prabhakaran V, and [CrossRef] [PubMed] | ||
| 153 | Fine A (1988) Evidence for an adrenal contribution to plasma digitalis-like factors. Clin Physiol Biochem 6:253–261. [CrossRef] [PubMed] | ||
| 154 | Gillis RA and Quest JA (1979) The role of the nervous system in the cardiovascular effects of digitalis. Pharmacol Rev 31:19 –97. [CrossRef] [PubMed] | ||
| 155 | Geering K (2005) Function of FXYD proteins, regulators of Na,K-ATPase. J Bioenerg Biomembr 37:387–392. [CrossRef] [PubMed] | ||
| 156 | Geering K (2006) FXYD proteins: new regulators of Na-K-ATPase. Am J Physiol Renal Physiol 290:F241–F250. Gerbi A, Barbey O, Raccah D, Coste T, Jamme I, Nouvelot A, Ouafik L, Lévy S, [CrossRef] [PubMed] | ||
| 157 | Vague P, and Maixent JM (1997) Alteration of Na,K-ATPase isoenzymes in diabetic cardiomyopathy: effect of dietary supplementation with fish oil (n-3 fatty acids) in rats. Diabetologia 40:496 –505. [CrossRef] [PubMed] | ||
| 158 | Ghoumari AM, Piochon C, Tomkiewicz C, Eychenne B, Levenes C, Dusart I, Schumacher M, and Baulieu EE (2006) Neuroprotective effect of mifepristone involves neuron depolarization. FASEB J 20:1377–1386. [CrossRef] [PubMed] | ||
| 159 | Gilson GJ, Graves SW, Qualls CR, and Curet LB (1997) Digoxin-like immunoreactive substance and sodium-potassium-adenosine triphosphatase inhibition in normal pregnancy: a longitudinal study. Obstet Gynecol 89:743–746. [CrossRef] [PubMed] | ||
| 160 | Glynn IM (1993) Annual Review prize lecture: ‘All hands to the sodium pump’. J Physiol 462:1–30. [CrossRef] [PubMed] | ||
| 161 | Goldstein I, Levy T, Galili D, Ovadia H, Yirmiya R, Rosen H, and Lichtstein D (2006) Involvement of Na+,K+-ATPase and endogenous digitalis-like compounds in depressive disorders. Biol Psychiatry 60:491– 499. [CrossRef] [PubMed] | ||
| 162 | Gonick HC, Ding Y, Vaziri ND, Bagrov AY, and Fedorova OV (1998) Simultaneous measurement of marinobufagenin, ouabain, and hypertension-associated protein in various disease states. Clin Exp Hypertens 20:617– 627. [CrossRef] [PubMed] | ||
| 163 | Goodlin RC (1988) Antidigoxin antibodies in eclampsia. N Engl J Med 318:518 –519. [CrossRef] [PubMed] | ||
| 164 | Göõz M, Tóth M, Vakkuri O, Göõz P, Smolka AJ, de Châtel R, and Szalay KS (2004) Endogenous ouabain-like factor (OLF) secretion is modulated by nicotinic mechanisms in rat adrenocortical cells. Life Sci 74:2111–2128. Goto A, Ishiguro T, Yamada K, Ishii M, Yoshioka M, Eguchi C, Shimora M, and [CrossRef] [PubMed] | ||
| 165 | Sugimoto T (1990) Isolation of a urinary digitalis-like factor indistinguishable from digoxin. Biochem Biophys Res Commun 173:1093–1101. [CrossRef] [PubMed] | ||
| 166 | Goto A, Yamada K, Ishii M, Sugimoto T, and Yoshioka M (1991a) Immunoreactivity of endogenous digitalis-like factors. Biochem Pharmacol 41:1261–1263. [CrossRef] [PubMed] | ||
| 167 | Goto A, Yamada K, Nagoshi H, Terano Y, and Omata M (1995) Stress-induced elevation of ouabainlike compound in rat plasma and adrenal. Hypertension 26: 1173–1176. immunoreactivity during surgical extirpation of pheochromocytoma. Hypertens Res 22:135–139. Komiyama Y, Nishimura N, Nishino N, Okuda K, Munakata M, Kosaka C, Masuda [CrossRef] [PubMed] | ||
| 168 | M, and Takahashi H (1998) Purification and characterization of ouabain-binding protein in human plasma. Clin Exp Hypertens 20:683– 690. Kotova O, Al-Khalili L, Talia S, Hooke C, Fedorova OV, Bagrov AY, and Chibalin AV (2006) Cardiotonic steroids stimulate glycogen synthesis in human skeletal muscle cells via a Srcand ERK1/2-dependent mechanism. J Biol Chem 281:20085–20094. [CrossRef] [PubMed] | ||
| 169 | Kramer HJ and Gonick HC (1974) Effect of extracellular volume expansion on renal Na-K-ATPase and cell metabolism. Nephron 12:281–296. Lam G, Johnson D, Robinson C, Saade G, Lewis D, Porter K, Humphrey R, Adair C, [CrossRef] [PubMed] | ||
| 170 | Buckalew V, and Danoff T (2008) Antepartum administration of a digoxin immune Fab (Digibind) improves renal function in patients with severe preeclampsia, in Proceedings of the XVIth Congress of the International Society for the Study of Hypertension in Pregnancy; 2008 Sept 20 –24; Washington DC. p 60, International Society for the Study of Hypertension in Pregnancy, Rotterdam, The Netherlands. [CrossRef] [PubMed] | ||
| 171 | Langer GA (1972) Effects of digitalis on myocardial ionic exchange. Circulation 46:180 –187. [CrossRef] [PubMed] | ||
| 172 | Laredo J, Hamilton BP, and Hamlyn JM (1994) Ouabain is secreted by bovine adrenocortical cells. Endocrinology 135:794 –797. [CrossRef] [PubMed] | ||
| 173 | Laredo J, Hamilton BP, and Hamlyn JM (1995) Secretion of endogenous ouabain from bovine adrenal cells: role of zona glomerulosa and zona fasciculata. Biochem Biophys Res Commun 212:487– 493. [CrossRef] [PubMed] | ||
| 174 | Laredo J, Shah JR, Lu ZR, Hamilton BP, and Hamlyn JM (1997) Angiotensin II stimulates secretion of endogenous ouabain from bovine adrenocortical cells via angiotensin type 2 receptors. Hypertension 29:401– 407. [CrossRef] [PubMed] | ||
| 175 | Larre I, Ponce A, Fiorentino R, Shoshani L, Contreras RG, and Cereijido M (2006) Contacts and cooperation between cells depend on the hormone ouabain. Proc Natl Acad Sci U S A 103:10911–10916. [CrossRef] [PubMed] | ||
| 176 | Lee CY and Burnett JC Jr (2007) Natriuretic peptides and therapeutic applications. Heart Fail Rev 12: 131–142. [CrossRef] [PubMed] | ||
| 177 | Leenen FH, Yuan B, and Huang BS (1999) Brain “ouabain” and angiotensin II contribute to cardiac dysfunction after myocardial infarction. Am J Physiol 277: H1786 –H1792. [CrossRef] [PubMed] | ||
| 178 | Lesney MS (2002) Flowers for the heart. Mod Drug Discov 5:46 – 48. [CrossRef] [PubMed] | ||
| 179 | Levenson R (1994) Isoforms of the Na,K-ATPase: family members in search of function. Rev Physiol Biochem Pharmacol 123:1– 45. Levinson PD, McGarvey ST, Carlisle CC, Eveloff SE, Herbert PN, and Millman RP (1993) Adiposity and cardiovascular risk factors in men with obstructive sleep apnea. Chest 103:1336 –1342. [CrossRef] [PubMed] | ||
| 180 | Li R, el-Mallakh RS, Harrison L, Changaris DG, and Levy RS (1997) Lithium prevents ouabain-induced behavioral changes toward an animal model for manic depression. Mol Chem Neuropathol 31:65–72. [CrossRef] [PubMed] | ||
| 181 | Liang M, Tian J, Liu L, Pierre S, Liu J, Shapiro J, and Xie ZJ (2007) Identification of a pool of non-pumping Na/K-ATPase. J Biol Chem 282:10585–10593. [CrossRef] [PubMed] | ||
| 182 | Lichtstein D, Gati I, Babila T, Haver E, and Katz U (1991) Effect of salt acclimation on digitalis-like compounds in the toad. Biochim Biophys Acta 1073:65– 68. [CrossRef] [PubMed] | ||
| 183 | Lichtstein D, Gati I, and Ovadia H (1993) Digitalis-like compounds in the toad Bufo viridis: interactions with plasma proteins. J Cardiovasc Pharmacol 22 (Suppl 2):S102–S105. Lichtstein D, Gati I, Samuelov S, Berson D, Rozenman Y, Landau L, and Deutsch J (1993) Identification of digitalis-like compounds in human cataractous lenses. Eur J Biochem 216:261–268. [CrossRef] [PubMed] | ||
| 184 | Lichtstein D, Steinitz M, Gati I, Samuelov S, Deutsch J, and Orly J (1998) Biosynthesis of digitalis-compound in rat adrenal cells: hydroxycholesterol as a precursor. Life Sci 62:2109 –2126. [CrossRef] [PubMed] | ||
| 185 | Lindenmayer GE (1976) Mechanism of action of digitalis glycosides at the subcellular level. Pharmacol Ther [B] 2:843– 861. [CrossRef] [PubMed] | ||
| 186 | Ling G (2007) History of the membrane (pump) theory of the living cell from its beginning in mid-19th century to its disproof 45 years ago—though still taught worldwide today as established truth. Physiol Chem Phys Med NMR 39:1– 67. [CrossRef] [PubMed] | ||
| 187 | Lingrel JB (1992) Na,K-ATPase: isoform structure, function, and expression. J Bioenerg Biomembr 24:263–270. [CrossRef] [PubMed] | ||
| 188 | Liu J, Kesiry R, Periyasamy SM, Malhotra D, Xie Z, and Shapiro JI (2004) Ouabain induces endocytosis of plasmalemmal Na/K-ATPase in LLC-PK1 cells by a clathrin-dependent mechanism. Kidney Int 66:227–241. [CrossRef] [PubMed] | ||
| 189 | Liu J, Liang M, Liu L, Malhotra D, Xie Z, and Shapiro JI (2005) Ouabain-induced endocytosis of the plasmalemmal Na/K-ATPase in LLC-PK1 cells requires caveolin-1. Kidney Int 67:1844 –1854. Liu J, Periyasamy SM, Gunning W, Fedorova OV, Bagrov AY, Malhotra D, Xie Z, and [CrossRef] [PubMed] | ||
| 190 | Shapiro JI (2002) Effects of cardiac glycosides on sodium pump expression and function in LLC-PK1 and MDCK cells. Kidney Int 62:2118 –2125. [CrossRef] [PubMed] | ||
| 191 | Liu J and Shapiro JI (2007) Regulation of sodium pump endocytosis by cardiotonic steroids: molecular mechanisms and physiological implications. Pathophysiology 14:171–181. [CrossRef] [PubMed] | ||
| 192 | Liu J, Tian J, Haas M, Shapiro JI, Askari A, and Xie Z (2000) Ouabain interaction with cardiac Na+/K+-ATPase initiates signal cascades independent of changes in intracellular Na+ and Ca2+ concentrations. J Biol Chem 275:27838 –27844. Liu L, Mohammadi K, Aynafshar B, Wang H, Li D, Liu J, Ivanov AV, Xie Z, and [CrossRef] [PubMed] | ||
| 193 | Askari A (2003) Role of caveolae in signal-transducing function of cardiac Na+/ K+-ATPase. Am J Physiol Cell Physiol 284:C1550 –C1560. [CrossRef] [PubMed] | ||
| 194 | Liu ZQ, Ma AQ, Zhang L, and Yang DY (1990) Intra-cellular electrolyte changes and levels of endogenous digoxin-like substance within the plasma in patients with congestive heart failure. Int J Cardiol 27:47–53. [CrossRef] [PubMed] | ||
| 195 | London GM and Parfrey PS (1997) Cardiac disease in chronic uremia: Pathogenesis. Adv Ren Replace Ther 4:194 –211. Lopatin DA, Ailamazian EK, Dmitrieva RI, Shpen VM, Fedorova OV, Doris PA, and [CrossRef] [PubMed] | ||
| 196 | Bagrov AY (1999) Circulating bufodienolide and cardenolide sodium pump inhibitors in preeclampsia. J Hypertens 17:1179 –1187. [CrossRef] [PubMed] | ||
| 197 | Loreaux EL, Kaul B, Lorenz JN, and Lingrel JB (2008) Ouabain-sensitive α1 Na,K- [CrossRef] [PubMed] | ||
| 198 | Hundal HS, Marette A, Mitsumoto Y, Ramlal T, Blostein R, and Klip A (1992) Insulin induces translocation of the α2 and β1 subunits of the Na+/K+-ATPase from intracellular compartments to the plasma membrane in mammalian skeletal muscle. J Biol Chem 267:5040 –5043. [CrossRef] [PubMed] | ||
| 199 | Itabashi A, Chan L, Shapiro JI, Cheung C, and Schrier RW (1987) Comparison of the natriuretic response to atriopeptin III and loop diuretic in the isolated perfused rat kidney. Clin Sci (Lond) 73:143–150. [CrossRef] [PubMed] | ||
| 200 | Iwamoto T (2006) Vascular Na+/Ca2+ exchanger: implications for the pathogenesis and therapy of salt-dependent hypertension. Am J Physiol Regul Integr Comp Physiol 290:R536 –R545. Iwamoto T, Kita S, Zhang J, Blaustein MP, Arai Y, Yoshida S, Wakimoto K, Komuro [CrossRef] [PubMed] | ||
| 201 | I, and Katsuragi T (2004) Salt-sensitive hypertension is triggered by Ca2+ entry via Na+/Ca2+ exchanger type-1 in vascular smooth muscle. Nat Med 10:1193– 1199. [CrossRef] [PubMed] | ||
| 202 | Jørgensen PL (1986) Structure, function and regulation of Na,K-ATPase in the kidney. Kidney Int 29:10 –20. [CrossRef] [PubMed] | ||
| 203 | Jørgensen PL (1990) Structure and molecular mechanisms of the Na,K-pump, in Monovalent Cations in Biological Systems (Pasternak CA, ed) pp 117–154, CRC, Boca Raton, FL. [CrossRef] [PubMed] | ||
| 204 | Juhaszova M and Blaustein MP (1997a) Distinct distribution of different Na+ pump α subunit isoforms in plasmalemma: physiological implications. Ann N Y Acad Sci 834:524 –536. [CrossRef] [PubMed] | ||
| 205 | Juhaszova M and Blaustein MP (1997b) Na+ pump low and high ouabain affinity α subunit isoforms are differently distributed in cells. Proc Natl Acad Sci U S A 94:1800 –1805. [CrossRef] [PubMed] | ||
| 206 | Kähönen M, Doris PA, Mäkynen H, and Pörsti I (1995) Plasma digoxin immunoreactivity and arterial potassium relaxation after quinapril therapy in hypertensive rats. J Pharmacol Exp Ther 275:832– 837. [CrossRef] [PubMed] | ||
| 207 | Kaminski K and Rechberger T (1991) Concentration of digoxin-like immunoreactive substance in patients with preeclampsia and its relation to severity of pregnancyinduced hypertension. Am J Obstet Gynecol 165:733–736. [CrossRef] [PubMed] | ||
| 208 | Kaplan MA, Bourgoignie JJ, Rosecan J, and Bricker NS (1974) The effects of the natriuretic factor from uremic urine on sodium transport, water and electrolyte content, and pyruvate oxidation by the isolated toad bladder. J Clin Invest 53: 1568 –1577. Kashkin VA, Zvartau EE, Fedorova OV, Bagrov YY, Lakatta EG, and Bagrov AY (2008) Endogenous bufadienolide mediates pressor response to ethanol withdrawal in rats. Eur Neuropsychopharmacol 18:74 –77. [CrossRef] [PubMed] | ||
| 209 | Kawazoe N, Aiuchi T, Masuda Y, Nakajo S, and Nakaya K (1999) Induction of apoptosis by bufalin in human tumor cells is associated with a change of intracellular concentration of Na+ ions. J Biochem 126:278 –286. [CrossRef] [PubMed] | ||
| 210 | Ke YS, Wang DG, Wang HG, and Yang SY (2004) Endoxin antagonist lessens myocardial ischemia reperfusion injury. Cardiovasc Drugs Ther 18:289 –293. [CrossRef] [PubMed] | ||
| 211 | Kelly RA (1986) Excretion of artifactual endogenous digitalis-like factors. Am J Physiol 251:H205–H209. [CrossRef] [PubMed] | ||
| 212 | Kelly RA and Smith TW (1992) Is ouabain the endogenous digitalis? Circulation 86:694 – 697. [CrossRef] [PubMed] | ||
| 213 | Kennedy D, Omran E, Periyasamy SM, Nadoor J, Priyadarshi A, Willey JC, Malhotra D, Xie Z, and Shapiro JI (2003) Effect of chronic renal failure on cardiac contractile function, calcium cycling, and gene expression of proteins important for calcium homeostasis in the rat. J Am Soc Nephrol 14:90 –97. Kennedy DJ, Elkareh J, Shidyak A, Shapiro AP, Smaili S, Mutgi K, Gupta S, Tian [CrossRef] [PubMed] | ||
| 214 | 2007 | J, Morgan E, Khouri S, et al | ( 2007 ) Partial nephrectomy as a model for uremic cardiomyopathy in the mouse. Am J Physiol Renal Physiol 294:F450 –F454. Kennedy DJ, Vetteth S, Periyasamy SM, Kanj M, Fedorova L, Khouri S, Kahaleh MB [CrossRef] [PubMed] |
| 215 | 2006a | Xie Z, Malhotra D, Kolodkin NI, Lakatta EG, et al | ( 2006a ) Central role for the cardiotonic steroid marinobufagenin in the pathogenesis of experimental uremic cardiomyopathy. Hypertension 47:488 – 495. Kennedy DJ, Vetteth S, Xie M, Periyasamy SM, Xie Z, Han C, Basrur V, Mutgi K, [CrossRef] [PubMed] |
| 216 | Fedorov V, Malhotra D, and Shapiro JI (2006b) Ouabain decreases sarco(endo)plasmic reticulum calcium ATPase activity in rat hearts by a process involving protein oxidation. Am J Physiol Heart Circ Physiol 291:H3003–H3011. [CrossRef] [PubMed] | ||
| 217 | Kieval RS, Butler VP Jr, Derguini F, Bruening RC, and Rosen MR (1988) Cellular electrophysiologic effects of vertebrate digitalis-like substances. J Am Coll Cardiol 11:637– 643. [CrossRef] [PubMed] | ||
| 218 | Klein M, Nejad NS, Lown B, Hagemeijer F, and Barr I (1971) Correlation of the electrical and mechanical changes in the dog heart during progressive digitalization. Circ Res 29:635– 645. Koga M, Ideishi M, Matsusaki M, Tashiro E, Kinoshita A, Ikeda M, Tanaka H, [CrossRef] [PubMed] | ||
| 219 | Shindo M, and Arakawa K (1992) Mild exercise decreases plasma endogenous digitalislike substance in hypertensive individuals. Hypertension 19: II231–II236. [CrossRef] [PubMed] | ||
| 220 | Kojima I, Yoshihara S, and Ogata E (1982) Involvement of endogenous digitalis-like substance in genesis of deoxycorticosterone-salt hypertension. Life Sci 30:1775– 1781. [CrossRef] [PubMed] | ||
| 221 | Kometiani P, Li J, Gnudi L, Kahn BB, Askari A, and Xie Z (1998) Multiple signal transduction pathways link Na+/K+-ATPase to growth-related genes in cardiac myocytes: the roles of Ras and mitogen-activated protein kinases. J Biol Chem 273:15249 –15256. Komiyama Y, Dong XH, Nishimura N, Masaki H, Yoshika M, Masuda M, and [CrossRef] [PubMed] | ||
| 222 | Takahashi H (2005) A novel endogenous digitalis, telocinobufagin, exhibits elevated plasma levels in patients with terminal renal failure. Clin Biochem 38:36 – 45. Komiyama Y, Kimura Y, Nishimura N, Hara K, Mori T, Okuda K, Munakata M, [CrossRef] [PubMed] | ||
| 223 | Masuda M, Murakami T, and Takahashi H (1997) Vasodepressor effects of exercise are accompanied by reduced circulating ouabainlike immunoreactivity and normalization of nitric oxide synthesis. Clin Exp Hypertens 19:363–372. Komiyama Y, Nishimura N, Munakata M, Mori T, Okuda K, Nishino N, Hirose S, [CrossRef] [PubMed] | ||
| 224 | Kosaka C, Masuda M, and Takahashi H (2001) Identification of endogenous ouabain in culture supernatant of PC12 cells. J Hypertens 19:229 –236. Komiyama Y, Nishimura N, Munakata M, Okuda K, Nishino N, Kosaka C, Masuda [CrossRef] [PubMed] | ||
| 225 | M, Mori T, Matsuda T, and Takahashi H (1999) Increases in plasma ouabainlike in vivo protective effect of atriopeptin III on ischemic acute renal failure. J Clin Invest 80:698 –705. Naruse K, Naruse M, Tanabe A, Yoshimoto T, Watanabe Y, Kurimoto F, Horiba N, [CrossRef] [PubMed] | ||
| 226 | Tamura M, Inagami T and Demura H (1994) Does plasma immunoreactive ouabain originate from the adrenal gland? Hypertension 23(1 Suppl):I102–I105. [CrossRef] [PubMed] | ||
| 227 | Naylor GJ, Dick DA, Dick EG, Le Poidevin D, and Whyte SF (1973) Erythrocyte membrane cation carrier in depressive illness. Psychol Med 3:502–508. [CrossRef] [PubMed] | ||
| 228 | Neuss M, Crow MT, Chesley A, and Lakatta EG (2001) Apoptosis in cardiac disease—what is it— how does it occur. Cardiovasc Drugs Ther 15:507–523. [CrossRef] [PubMed] | ||
| 229 | Newman RA, Yang P, Pawlus AD, and Block KI (2008) Cardiac glycosides as novel cancer therapeutic agents. Mol Interv 8:36 – 49. [CrossRef] [PubMed] | ||
| 230 | Nesher M, Shpolansky U, Rosen H, and Lichtstein D (2007) The digitalis-like steroid hormones: new mechanisms of action and biological significance. Life Sci 80:2093– 2107. [CrossRef] [PubMed] | ||
| 231 | Nguyen AN, Wallace DP, and Blanco G (2007) Ouabain binds with high affinity to the Na,K-ATPase in human polycystic kidney cells and induces extracellular signal-regulated kinase activation and cell proliferation. J Am Soc Nephrol 18:46 – 57. [CrossRef] [PubMed] | ||
| 232 | Nhamburo PT, Salafsky BP, Hoffman PL, and Tabakoff B (1986) Effects of shortchain alcohols and norepinephrine on brain Na,K-ATPase activity. Biochem Pharmacol 35:1987–1992. [CrossRef] [PubMed] | ||
| 233 | Nhamburo PT, Salafsky BP, Tabakoff B, and Hoffman PL (1987) Effects of ethanol on ouabain inhibition of mouse brain Na,K-ATPase activity. Biochem Pharmacol 36:2027–2033. [CrossRef] [PubMed] | ||
| 234 | Nicholls MG, Lewis LK, Yandle TG, Lord G, McKinnon W, and Hilton PJ (2009) Ouabain, a circulating hormone secreted by the adrenals, is pivotal in cardiovascular disease: fact or fantasy? J Hypertens 27:3– 8. [CrossRef] [PubMed] | ||
| 235 | Numazawa S, Honma Y, Yamamoto T, Yoshida T, and Kuroiwa Y (1995) A cardiotonic steroid bufalin-like factor in human plasma induces leukemia cell differentiation. Leuk Res 19:945–953. [CrossRef] [PubMed] | ||
| 236 | Numazawa S, Shinoki MA, Ito H, Yoshida T, and Kuroiwa Y (1994) Involvement of Na,K-ATPase inhibition in K562 cell differentiation induced by bufalin. J Cell Physiol 160:113–120. Oda M, Kurosawa M, Numazawa S, Tanaka S, Akizawa T, Ito K, Maeda M, and [CrossRef] [PubMed] | ||
| 237 | Yoshida T (2001) Determination of bufalin-like immunoreactivity in serum of humans and rats by time-resolved fluoroimmunoassay for using a monoclonal antibody. Life Sci 68:1107–1117. [CrossRef] [PubMed] | ||
| 238 | Orlov SN and Hamet P (2004) Apoptosis vs. oncosis: role of cell volume and intracellular monovalent cations. Adv Exp Med Biol 559:219 –233. [CrossRef] [PubMed] | ||
| 239 | Orlov SN and Hamet P (2006) The death of cardiotonic steroid-treated cells: evidence of Na+i,K+i-independent H+i-sensitive signalling. Acta Physiol (Oxf) 187:231–240. [CrossRef] [PubMed] | ||
| 240 | Orlov SN and Mongin AA (2007) Salt-sensing mechanisms in blood pressure regulation and hypertension. Am J Physiol Heart Circ Physiol 293:H2039 –H2053. Orlov SN, Thorin-Trescases N, Pchejetski D, Taurin S, Farhat N, Tremblay J, Thorin [CrossRef] [PubMed] | ||
| 241 | E, and Hamet P (2004) Na+/K+ pump and endothelial cell survival: [Na+]i/[K+]iindependent necrosis triggered by ouabain, and protection against apoptosis mediated by elevation of [Na+]i. Pflugers Arch 448:335–345. [CrossRef] [PubMed] | ||
| 242 | Overbeck HW, Pamnani MB, Akera T, Brody TM, and Haddy FJ (1976) Depressed function of a ouabain-sensitive sodium-potassium pump in blood vessels from renal hypertensive dogs. Circ Res 38(6 Suppl 2):48 –52. Oweis S, Wu L, Kiela PR, Zhao H, Malhotra D, Ghishan FK, Xie Z, Shapiro JI, and [CrossRef] [PubMed] | ||
| 243 | Liu J (2006) Cardiac glycoside downregulates NHE3 activity and expression in LLC-PK1 cells. Am J Physiol Renal Physiol 290:F997–F1008. Paci A, Marrone O, Lenzi S, Prontera C, Nicolini G, Ciabatti G, Ghione S, and [CrossRef] [PubMed] | ||
| 244 | Bonsignore G (2000) Endogenous digitalis-like factors in obstructive sleep apnea. Hypertens Res 23 (Suppl):S87–S91. [CrossRef] [PubMed] | ||
| 245 | Padilha AS, Peçanha FM, Vassallo DV, Alonso MJ, and Salaices M (2008) Ouabain treatment changes the role of endothelial factors in rat resistance arteries. Eur J Pharmacol 600:110 –116. [CrossRef] [PubMed] | ||
| 246 | Pamnani MB, Whitehorn WV, Clough DL, and Haddy FJ (1990) Effects of canrenone on blood pressure in rats with reduced renal mass. Am J Hypertens 3:188 –195. [CrossRef] [PubMed] | ||
| 247 | Parhami-Seren B, Haberly R, Margolies MN, and Haupert GT Jr (2002) Ouabainbinding protein(s) from human plasma. Hypertension 40:220 –228. Periyasamy SM, Chen J, Cooney D, Carter P, Omran E, Tian J, Priyadarshi S, [CrossRef] [PubMed] | ||
| 248 | 2001 | Bagrov A, Fedorova O, Malhotra D, et al | ( 2001 ) Effects of uremic serum on isolated cardiac myocyte calcium cycling and contractile function. Kidney Int 60:2367–2376. Periyasamy SM, Liu J, Tanta F, Kabak B, Wakefield B, Malhotra D, Kennedy DJ, [CrossRef] [PubMed] |
| 249 | 2005 | Nadoor A, Fedorova OV, Gunning W, et al | ( 2005 ) Salt loading induces redistribution of the plasmalemmal Na/K-ATPase in proximal tubule cells. Kidney Int 67:1868 –1877. [CrossRef] [PubMed] |
| 250 | Perrin A, Brasmes B, Chambaz EM, and Defaye G (1997) Bovine adrenocortical cells in culture synthesize an ouabain-like compound. Mol Cell Endocrinol 126:7–15. [CrossRef] [PubMed] | ||
| 251 | Peterson GL, Ewing RD, Hootman SR, and Conte FP (1978) Large scale partial purification and molecular and kinetic properties of the (Na+K+)-activated adenosine triphosphatase from Artemia salina nauplii. J Biol Chem 253:4762– 4770. Pierdomenico SD, Bucci A, Manunta P, Rivera R, Ferrandi M, Hamlyn JM, Lapenna [CrossRef] [PubMed] | ||
| 252 | D, Cuccurullo F, and Mezzetti A (2001) Endogenous ouabain and hemodynamic and left ventricular geometric patterns in essential hypertension. Am J Hypertens 14:44 –50. [CrossRef] [PubMed] | ||
| 253 | Pierre SV, Sottejeau Y, Gourbeau JM, Sánchez G, Shidyak A, and Blanco G (2008) Isoform-specificity of Na,K-ATPase-mediated ouabain signaling. Am J Physiol Renal Physiol 294:F859 –F866. [CrossRef] [PubMed] | ||
| 254 | Pierre SV and Xie Z (2006) The Na,K-ATPase receptor complex: its organization and membership. Cell Biochem Biophys 46:303–316. [CrossRef] [PubMed] | ||
| 255 | Pleasants RA, Gadsden RH Sr, McCormack JP, Piveral K, and Sawyer WT (1986) Interference of digoxin-like immunoreactive substances with three digoxin immunoassays in patients with various degrees of renal function. Clin Pharm 5:810 – 816. [CrossRef] [PubMed] | ||
| 256 | Porto AM and Gros EG (1970) Biosynthesis of animal and plant bufadienolides: ATPase enhances natriuretic response to saline load. J Am Soc Nephrol 19:1947– 1954. Lorenz JN, Loreaux EL, Dostanic-Larson I, Lasko V, Schnetzer JR, Paul RJ, and [CrossRef] [PubMed] | ||
| 257 | Lingrel JB (2008) ACTH-induced hypertension is dependent on the ouabainbinding site of the α2-Na+-K+-ATPase subunit. Am J Physiol Heart Circ Physiol 295:H273–H280. Ludens JH, Clark MA, DuCharme DW, Harris DW, Lutzke BS, Mandel F, Mathews [CrossRef] [PubMed] | ||
| 258 | WR, Sutter DM, and Hamlyn JM (1991) Purification of an endogenous digitalislike factor from human plasma for structural analysis. Hypertension 17:923–929. [CrossRef] [PubMed] | ||
| 259 | Ludens JH, Clark MA, Kolbasa KP and Hamlyn JM (1993) Digitalis-like factor and ouabain-like compound in plasma of volume-expanded dogs. J Cardiovasc Pharmacol 22 (Suppl 2):S38 –S41. [CrossRef] [PubMed] | ||
| 260 | Lyttle T, Goldstein D, and Gartz J (1996) Bufo toads and bufotenine: fact and fiction surrounding an alleged psychedelic. J Psychoactive Drugs 28:267–290. [CrossRef] [PubMed] | ||
| 261 | Lytton J, Lin JC, and Guidotti G (1985) Identification of two molecular forms of (Na,K)-ATPase in rat adipocytes. Relation to insulin stimulation of the enzyme. J Biol Chem 260:1177–1184. [CrossRef] [PubMed] | ||
| 262 | Maixent JM and Lelièvre LG (1987) Differential inactivation of inotropic and toxic digitalis receptors in ischemic dog heart: molecular basis of the deleterious effects of digitalis. J Biol Chem 262:12458 –12462. [CrossRef] [PubMed] | ||
| 263 | Manunta P, Ferrandi M, Bianchi G, and Hamlyn JM (2009) Endogenous ouabain in cardiovascular function and disease. J Hypertens 27:9 –18. [CrossRef] [PubMed] | ||
| 264 | Manunta P, Hamilton BP, and Hamlyn JM (2006) Salt intake and depletion increase circulating levels of endogenous ouabain in normal men. Am J Physiol Regul Integr Comp Physiol 290:R553–R559. Manunta P, Maillard M, Tantardini C, Simonini M, Lanzani C, Citterio L, Stella P, [CrossRef] [PubMed] | ||
| 265 | 2008 | Casamassima N, Burnier M, Hamlyn JM, et al | ( 2008 ) Relationships among endogenous ouabain, α-adducin polymorphisms and renal sodium handling in primary hypertension. J Hypertens 26:914 –920. Manunta P, Messaggio E, Ballabeni C, Sciarrone MT, Lanzani C, Ferrandi M, [CrossRef] [PubMed] |
| 266 | Hamlyn JM, Cusi D, Galletti F, and Bianchi G (2001) Plasma ouabain-like factor during acute and chronic changes in sodium balance in essential hypertension. Hypertension 38:198 –203. Manunta P, Stella P, Rivera R, Ciurlino D, Cusi D, Ferrandi M, Hamlyn JM, and [CrossRef] [PubMed] | ||
| 267 | Bianchi G (1999) Left ventricular mass, stroke volume, and ouabain-like factor in essential hypertension. Hypertension 34:450 – 456. [CrossRef] [PubMed] | ||
| 268 | Markou A, Kosten TR, and Koob GF (1998) Neurobiological similarities in depression and drug dependence: a self-medication hypothesis. Neuropsychopharmacology 18:135–174. [CrossRef] [PubMed] | ||
| 269 | Marks MJ and Seeds NW (1978) A heterogeneous ouabain-ATPase interaction in mouse brain. Life Sci 23:2735–2744. [CrossRef] [PubMed] | ||
| 270 | Martinka E, Galajada P, Ochodnicky M, Lichardus B, Straka S, and Mokan M (1997) Endogenous digoxinlike immunoreactivity and diabetes mellitus: facts and hypotheses. Med Hypotheses 49:271–275. [CrossRef] [PubMed] | ||
| 271 | Masilamani S, Castro L, and Baylis C (1994) Pregnant rats are refractory to the natriuretic actions of ANP. Am J Physiol 267:R1611–R1616. [CrossRef] [PubMed] | ||
| 272 | Masuda Y, Kawazoe N, Nakajo S, Yoshida T, Kuroiwa Y, and Nakaya K (1995) Bufalin induces apoptosis and influences the expression of apoptosis-related genes in human leukemia cells. Leuk Res 19:549 –556. [CrossRef] [PubMed] | ||
| 273 | McDonough AA, Wang J, and Farley RA (1995) Significance of sodium pump isoforms in digitalis therapy. J Mol Cell Cardiol 27:1001–1009. [CrossRef] [PubMed] | ||
| 274 | McGrail KM, Phillips JM, and Sweadner KJ (1991) Immunofluorescent localization of three Na,K-ATPase isozymes in the rat central nervous system: both neurons and glia can express more than one Na,K-ATPase. J Neurosci 11:381–391. [CrossRef] [PubMed] | ||
| 275 | Meneton P, Jeunemaitre X, de Wardener HE, and MacGregor GA (2005) Links between dietary salt intake, renal salt handling, blood pressure, and cardiovascular diseases. Physiol Rev 85:679 –715. [CrossRef] [PubMed] | ||
| 276 | Menezes JC, Troster EJ, and Dichtchekenian V (2003) Digoxin antibody decreases natriuresis and diuresis in cerebral hemorrhage. Intensive Care Med 29:2291– 2296. [CrossRef] [PubMed] | ||
| 277 | Meyer K and Linde H (1971) Collection of toad venoms and chemistry of the toad venom steroids, in Venomous Animals and Their Venoms (Bucherl W and Buckley E eds), Academic Press, London. [CrossRef] [PubMed] | ||
| 278 | Middleton RJ, Parfrey PS, and Foley RN (2001) Left ventricular hypertrophy in the renal patient. J Am Soc Nephrol 12:1079 –1084. Mijatovic T, De Nève N, Gailly P, Mathieu V, Haibe-Kains B, Bontempi G, Lapeira [CrossRef] [PubMed] | ||
| 279 | J, Decaestecker C, Facchini V, and Kiss R (2008) Nucleolus and c-Myc: potential targets of cardenolide-mediated antitumor activity. Mol Cancer Ther 7:1285–1296. [CrossRef] [PubMed] | ||
| 280 | Mijatovic T, Van Quaquebeke E, Delest B, Debeir O, Darro F, and Kiss R (2007) Cardiotonic steroids on the road to anti-cancer therapy. Biochim Biophys Acta 1776:32–57. [CrossRef] [PubMed] | ||
| 281 | Miller WL (2008) Steroidogenic enzymes. Endocr Dev 13:1–18. Mobasheri A, Avila J, Cózar-Castellano I, Brownleader MD, Trevan M, Francis MJ, [CrossRef] [PubMed] | ||
| 282 | Lamb JF, and Martín-Vasallo P (2000) Na+,K+-ATPase isozyme diversity; comparative biochemistry and physiological implications of novel functional interactions. Biosci Rep 20:51–91. [CrossRef] [PubMed] | ||
| 283 | Mohmand B, Malhotra DK, and Shapiro JI (2005) Uremic cardiomyopathy: role of circulating digitalis like substances. Front Biosci 10:2036 –2044. [CrossRef] [PubMed] | ||
| 284 | Morise T, Okamoto S, Takasaki H, Ikeda M, Takeda R, Kiuti F, and Tuda Y (1988) Biological activity of partially purified digitalis-like substance and Na-K-ATPase inhibitor in rats. Jpn Circ J 52:1309 –1316. Müller-Ehmsen J, Juvvadi P, Thompson CB, Tumyan L, Croyle M, Lingrel JB, [CrossRef] [PubMed] | ||
| 285 | Schwinger RH, McDonough AA, and Farley RA (2001) Ouabain and substrate affinities of human Na+-K+-ATPase alpha(1)beta(1), alpha(2)beta(1), and alpha(3)beta(1) when expressed separately in yeast cells. Am J Physiol Cell Physiol 281:C1355–C1364. Murrell JR, Randall JD, Rosoff J, Zhao JL, Jensen RV, Gullans SR, and Haupert GT Jr (2005) Endogenous ouabain: upregulation of steroidogenic genes in hypertensive hypothalamus but not adrenal. Circulation 112:1301–1308. [CrossRef] [PubMed] | ||
| 286 | Nakamoto M, Shapiro JI, Shanley PF, Chan L, and Schrier RW (1987) In vitro and (1995) Ouabain-inhibiting activity of aldosterone antagonists. Steroids 60:110 – 113. [CrossRef] [PubMed] | ||
| 287 | Shah JR, Laredo J, Hamilton BP, and Hamlyn JM (1998) Different signaling pathways mediate stimulated secretions of endogenous ouabain and aldosterone from bovine adrenocortical cells. Hypertension 31:463– 468. [CrossRef] [PubMed] | ||
| 288 | Shah JR, Laredo J, Hamilton BP, and Hamlyn JM (1999) Effects of angiotensin II on sodium potassium pumps, endogenous ouabain, and aldosterone in bovine zona glomerulosa cells. Hypertension 33:373–377. Shamraj OI, Grupp IL, Grupp G, Melvin D, Gradoux N, Kremers W, Lingrel JB, and [CrossRef] [PubMed] | ||
| 289 | De Pover A (1993) Characterisation of Na/K-ATPase, its isoforms, and the inotropic response to ouabain in isolated failing human hearts. Cardiovasc Res 27:2229 – 2237. [CrossRef] [PubMed] | ||
| 290 | Shamraj OI and Lingrel JB (1994) A putative fourth Na+,K+-ATPase α-subunit gene is expressed in testis. Proc Natl Acad Sci U S A 91:12952–12956. [CrossRef] [PubMed] | ||
| 291 | Sheiner LB, Rosenberg B, Marathe VV, and Peck C (1974) Differences in serum digoxin concentrations between outpatients and inpatients: an effect of compliance? Clin Pharmacol Ther 15:239 –246. [CrossRef] [PubMed] | ||
| 292 | Shrivastav P, Gill DS, D’Souza V, O’Brien PM, and Dandona P (1988) Secretion of atrial natriuretic peptide and digoxin-like immunoreactive substance during pregnancy. Clin Chem 34:977–980. [CrossRef] [PubMed] | ||
| 293 | Shull GE, Greeb J, and Lingrel JB (1986) Molecular cloning of three distinct forms of the Na+,K+-ATPase subunit from rat brain. Biochemistry 25:8125– 8132. [CrossRef] [PubMed] | ||
| 294 | Shull GE, Schwartz A, and Lingrel JB (1985) Amino-acid sequence of the catalytic subunit of the (Na+ + K+)ATPase deduced from a complementary DNA. Nature 316:61– 695. [CrossRef] [PubMed] | ||
| 295 | Sibai B, Dekker G, and Kupferminc M (2005) Pre-eclampsia. Lancet 365:785–799. [CrossRef] [PubMed] | ||
| 296 | Sich B, Kirch U, Tepel M, Zidek W, and Schoner W (1996) Pulse pressure correlates in humans with a proscillaridin A immunoreactive compound. Hypertension 27: 1073–1078. [CrossRef] [PubMed] | ||
| 297 | Siperstein MD, Murray AW, and Titus E (1957) Biosynthesis of cardiotonic sterols from cholesterol in the toad, Bufo marinus. Arch Biochem Biophys 67:154 –160. [CrossRef] [PubMed] | ||
| 298 | Skou JC (1957) The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim Biophys Acta 23:394 – 401. [CrossRef] [PubMed] | ||
| 299 | Skou JC and Esmann M (1992) The Na,K-ATPase. J Bioenerg Biomembr 24:249 – 261. [CrossRef] [PubMed] | ||
| 300 | Smith TW and Haber E (1973) Digitalis. I. N Engl J Med 289:945–952. [CrossRef] [PubMed] | ||
| 301 | Sophocleous A, Elmatzoglou I, and Souvatzoglou A (2003) Circulating endogenous digitalis-like factor(s) (EDLF) in man is derived from the adrenals and its secretion is ACTH-dependent. J Endocrinol Invest 26:668 – 674. [CrossRef] [PubMed] | ||
| 302 | Söderholm M, Hebert H, Skriver E, and Maunsbach AB (1988) Assembly of twodimensional membrane crystal of Na,K-ATPase. J Ultrastruct Mol Struct Res 99:234 –243. Staessen JA, Kuznetsova T, Acceto R, Bacchieri A, Brand E, Burnier M, Celis H, [CrossRef] [PubMed] | ||
| 303 | 2005 | Citterio L, de Leeuw PW, Filipovský J, et al | ( 2005 ) OASIS-HT: design of a pharmacogenomic dose-finding study. Pharmacogenomics 6:755–775. [CrossRef] [PubMed] |
| 304 | Stamler J, Rose G, Elliott P, Dyer A, Marmot M, Kesteloot H, and Stamler R (1991) Findings of the International Cooperative INTERSALT Study. Hypertension 17 (Suppl 1):I9 –I15. [CrossRef] [PubMed] | ||
| 305 | Stenkvist B, Pengtsson E, Dahlquist B, Eriksson O, Jarkrans T, and Nordin B (1982) Cardiac glycosides and breast cancer, revisited. N Engl J Med 306:484. Stoklosa T, Poplawski T, Koptyra M, Nieborowska-Skorska M, Basak G, Slupianek [CrossRef] [PubMed] | ||
| 306 | 2008 | A, Rayevskaya M, Seferynska I, Herrera L, Blasiak J, et al | ( 2008 ) BCR/ABL inhibits mismatch repair to protect from apoptosis and induce point mutations. Cancer Res 68:2576 –2580. Straub RH, Hall C, Krämer BK, Elbracht R, Palitzsch KD, Lang B, and Schölmerich J (1996) Atrial natriuretic factor and digoxin-like immunoreactive factor in diabetic patients: their interrelation and the influence of the autonomic nervous system. J Clin Endocrinol Metab 81:3385–3389. Sverdlov ED, Monastyrskaya GS, Broude NE, Ushkaryov YuA, Allikmets RL, [CrossRef] [PubMed] |
| 307 | Melkov AM, Smirnov YuV, Malyshev IV, Dulobova IE, and Petrukhin KE (1987) The family of human Na+,K+-ATPase genes: no less than five genes and/or pseudogenes related to the subunit. FEBS Lett 217:275–278. [CrossRef] [PubMed] | ||
| 308 | Sweadner KJ (1979) Two molecular forms of (Na+ + K+)-stimulated ATPase in brain: separation, and difference in affinity for strophanthidin. J Biol Chem 254:6060 – 6067. [CrossRef] [PubMed] | ||
| 309 | Sweadner KJ (1989) Isozymes of the Na+/K+-ATPase. Biochim Biophys Acta 988: 185–220. [CrossRef] [PubMed] | ||
| 310 | Sweadner KJ and Rael E (2000) The FXYD gene family of small ion transport regulators or channels: cDNA sequence, protein signature sequence, and expression. Genomics 68:41–56. [CrossRef] [PubMed] | ||
| 311 | Szent-Gyorgyi A (1953) Chemical Physiology of Contraction in Body and Heart Muscle. Academic Press, New York. Takahashi H, Iyoda I, Takeda K, Sasaki S, Okajima H, Yamasaki H, Yoshimura M, and Ijichi H (1984) Centrally-induced vasopressor responses to sodium-potassium adenosine triphosphatase inhibitor, ouabain, may be mediated via angiotensin II in the anteroventral third ventricle in the brain. Jpn Circ J 48:1243–1250. Takahashi H, Matsusawa M, Ikegaki I, Suga K, Nishimura M, Yoshimura M, [CrossRef] [PubMed] | ||
| 312 | Yamada H, and Sano Y (1988) Digitalis-like substance is produced in the hypothalamus but not in the adrenal gland in rats. J Hypertens Suppl 6:S345–S457. [CrossRef] [PubMed] | ||
| 313 | Tao QF, Soszynski PA, Hollenberg NK, and Graves SW (1996) Specificity of the volume-sensitive sodium pump inhibitor isolated from human peritoneal dialysate in chronic renal failure. Kidney Int 49:420 – 429. [CrossRef] [PubMed] | ||
| 314 | Therien AG and Blostein R (2000) Mechanisms of sodium pump regulation. Am J Physiol Cell Physiol 279:C541–C566. Tian J, Cai T, Yuan Z, Wang H, Liu L, Haas M, Maksimova E, Huang XY, and Xie ZJ (2006) Binding of Src to Na+/K+-ATPase forms a functional signaling complex. Mol Biol Cell 17:317–326. [CrossRef] [PubMed] | ||
| 315 | Tian J, Liu J, Garlid KD, Shapiro JI, and Xie Z (2003) Involvement of mitogenactivated protein kinases and reactive oxygen species in the inotropic action of ouabain on cardiac myocytes: a potential role for mitochondrial KATP channels. Mol Cell Biochem 242:181–187. parallel experiments with pregn-5-en-β-ol-20-one-20C in Scilla maritima and Bufo paracnemis. Experientia 26:11. [CrossRef] [PubMed] | ||
| 316 | Porto AM and Gros EG (1971) Biosynthesis of the bufadienolide marinobufagin in the toad Bufo paracnemis from cholesterol-20-14C. Experientia 27:506. [CrossRef] [PubMed] | ||
| 317 | Post RL, Hegyvary C, and Kume S (1972) Activation by adenosine triphosphate in the phosphorylation kinetics of sodium and potassium ion transport adenosine triphosphatase. J Biol Chem 247:6530 – 6540. [CrossRef] [PubMed] | ||
| 318 | Poston L, Morris JF, Wolfe CD, and Hilton PJ (1989) Serum digoxin-like substances in pregnancy-induced hypertension. Clin Sci (Lond) 77:189 –194. Priyadarshi S, Valentine B, Han C, Fedorova OV, Bagrov AY, Liu J, Periyasamy SM, [CrossRef] [PubMed] | ||
| 319 | 2003 | Kennedy D, Malhotra D, Xie Z, et al | ( 2003 ) Effect of green tea extract on cardiac hypertrophy following 5/6 nephrectomy in the rat. Kidney Int 63:1785–1790. [CrossRef] [PubMed] |
| 320 | Qazzaz HM, Cao Z, Bolanowski DD, Clark BJ, and Valdes R Jr (2004) De novo biosynthesis and radiolabeling of mammalian digitalis-like factors. Clin Chem 50:612– 620. [CrossRef] [PubMed] | ||
| 321 | Qazzaz HM, Goudy SL, and Valdes R Jr (1996) Deglycosylated products of endogenous digoxin-like immunoreactive factor in mammalian tissue. J Biol Chem 271: 8731– 8737. [CrossRef] [PubMed] | ||
| 322 | Rein H (1949) Uber ein Regulaionssystem “Mils-Leber” fur den oxydativen stiffwechsel der Korperwebe und besonders des Herzens. Naturwissenschaften 8:233–239. [CrossRef] [PubMed] | ||
| 323 | Repke KR (1972) Biochemical principles of the development of new cardiac drugs of the digitalis type. Pharmazie 27:693–701. [CrossRef] [PubMed] | ||
| 324 | Ritz E (1994) Presentation of the 1993 Jean Hamburger Award to Hugh E. de Wardener by Eberhard Ritz. Kidney Int Suppl 44:S114 –S115. [CrossRef] [PubMed] | ||
| 325 | Ritz E (1996) The history of salt—aspects of interest to the nephrologist. Nephrol Dial Transplant 11:969 –975. [CrossRef] [PubMed] | ||
| 326 | Ritz E, Dikow R, Morath C, and Schwenger V (2006) Salt—a potential ‘uremic toxin’? Blood Purif 24:63– 66. [CrossRef] [PubMed] | ||
| 327 | Rodrigues-Mascarenhas S, dos Santos NF, and Rumjanek VM (2006) Synergistic effect between ouabain and glucocorticoids for the induction of thymic atrophy. Biosci Rep 26:159 –169. Rossoni LV, Salaices M, Miguel M, Briones AM, Barker LA, Vassallo DV, and Alonso MJ (2002) Ouabain-induced hypertension is accompanied by increases in endothelial vasodilator factors. Am J Physiol Heart Circ Physiol 283:H2110 –H2118. [CrossRef] [PubMed] | ||
| 328 | Rossoni LV, Xavier FE, Moreira CM, Falcochio D, Amanso AM, Tanoue CU, Carvalho CR, and Vassallo DV (2006) Ouabain-induced hypertension enhances left ventricular contractility in rats. Life Sci 79:1537–1545. [CrossRef] [PubMed] | ||
| 329 | Rubattu S and Volpe M (2001) The atrial natriuretic peptide: a changing view. J Hypertens 19:1923–1931. [CrossRef] [PubMed] | ||
| 330 | Ruktanonchai DJ, el-Mallakh RS, Li R, and Levy RS (1998) Persistent hyperreactivity following a single intracerebroventricular dose of ouabain. Physiol Behav 63:403– 406. [CrossRef] [PubMed] | ||
| 331 | Russo JJ and Sweadner KJ (1993) Na+-K+-ATPase subunit isoform pattern modification by mitogenic insulin concentration in 3T3–L1 preadipocytes. Am J Physiol 264:C311–C316. [CrossRef] [PubMed] | ||
| 332 | Rybakowski JK and Lehmann W (1994) Decreased activity of erythrocyte membrane ATPases in depression and schizophrenia. Neuropsychobiology 30:11–14. [CrossRef] [PubMed] | ||
| 333 | Scavone C, Scanlon C, McKee M, and Nathanson JA (1995) Atrial natriuretic peptide modulates sodium and potassium-activated adenosine triphosphatase through a mechanism involving cyclic GMP and cyclic GMP-dependent protein kinase. J Pharmacol Exp Ther 272:1036 –1043. [CrossRef] [PubMed] | ||
| 334 | Schmidt RW, Bourgoignie JJ, and Bricker NS (1974) On the adaptation in sodium excretion in chronic uremia: the effects of “proportional reduction” of sodium intake. J Clin Invest 53:1736 –1741. Schneider R, Wray V, Nimtz M, Lehmann WD, Kirch U, Antolovic R, and Schoner W (1998) Bovine adrenals contain, in addition to ouabain, a second inhibitor of the sodium pump. J Biol Chem 273:784 –792. [CrossRef] [PubMed] | ||
| 335 | Schoner W (1992) Endogenous digitalis-like factors. Clin Exp Hypertens A 14:767– 814. [CrossRef] [PubMed] | ||
| 336 | Schoner W and Scheiner-Bobis G (2007) Endogenous and exogenous cardiac glycosides: their roles in hypertension, salt metabolism, and cell growth. Am J Physiol Cell Physiol 293:C509 –C536. [CrossRef] [PubMed] | ||
| 337 | Schreiber V, Kölbel F, Stĕpán J, Gregorová I, and Pribyl T (1981a) Digoxin-like immunoreactivity in the serum of rats with cardiac overload. J Mol Cell Cardiol 13:107–110. [CrossRef] [PubMed] | ||
| 338 | Schreiber V, Stĕpán J, Gregorová I, and Krejcíková J (1981b) Crossed digoxin immunoreactivity in chromatographic fractions of rat adrenal extract. Biochem Pharmacol 30:805– 806. [CrossRef] [PubMed] | ||
| 339 | Schrier RW (1974) Effects of adrenergic nervous system and catecholamines on systemic and renal hemodynamics, sodium and water excretion and renin secretion. Kidney Int 6:291–306. [CrossRef] [PubMed] | ||
| 340 | Schrier RW and Abraham WT (1999) Hormones and hemodynamics in heart failure. N Engl J Med 341:577–585. [CrossRef] [PubMed] | ||
| 341 | Schrier RW, McDonald KM, Marshall RA, and Lauler DP (1968a) Absence of natriuretic response to acute hypotonic intravascular volume expansion in dogs. Clin Sci 34:57–72. [CrossRef] [PubMed] | ||
| 342 | Schrier RW, Verroust PJ, Jones JJ, Fabian M, Lee J, and De Wardener HE (1968b) Effect of isotonic saline infusion and acute haemorrhage on plasma oxytocin and vasopressin concentrations in dogs. Clin Sci 35:433– 443. Schwinger RH, Wang J, Frank K, Müller-Ehmsen J, Brixius K, McDonough AA, and [CrossRef] [PubMed] | ||
| 343 | Erdmann E (1999) Reduced sodium pump α1, α3 and α1-isoform protein levels and Na+,K+-ATPase activity but unchanged Na+-Ca2+ exchanger protein levels in human heart failure. Circulation 99:2105–2512. [CrossRef] [PubMed] | ||
| 344 | Selye H, Mécs I, and Tamura T (1969) Effect of spironolactone and norbolethone on the toxicity of digitalis compounds in the rat. Br J Pharmacol 37:485– 488. Semplicini A, Buzzaccarini F, Ceolotto G, Marzola M, Mozzato MG, Giusto M, [CrossRef] [PubMed] | ||
| 345 | Campagnolo M, Simonella C, and Pessina AC (1993) Effects of canrenoate on red cell sodium transport and calf flow in essential hypertension. Am J Hypertens 6:295–301. Semplicini A, Serena L, Valle R, Ceolotto G, Felice M, Fontebasso A, and Pessina AC [CrossRef] [PubMed] | ||
| 346 | Wasserstrom JA and Aistrup GL (2005) Digitalis: new actions for an old drug. Am J Physiol Heart Circ Physiol 289:H1781–H1793. [CrossRef] [PubMed] | ||
| 347 | Watabe M, Masuda Y, Nakajo S, Yoshida T, Kuroiwa Y, and Nakaya K (1996) The cooperative interaction of two different signaling pathways in response to bufalin induces apoptosis in human leukemia U937 cells. J Biol Chem 271:14067–14072. [CrossRef] [PubMed] | ||
| 348 | Weidemann H (2005) Na/K-ATPase, endogenous digitalis like compounds and cancer development—a hypothesis. Front Biosci 10:2165–2176. Weidemann H, Salomon N, Avnit-Sagi T, Weidenfeld J, Rosen H, and Lichtstein D (2004) Diverse effects of stress and additional adrenocorticotropic hormone on digitalis-like compounds in normal and nude mice. J Neuroendocrinol 16:458 – 463. [CrossRef] [PubMed] | ||
| 349 | Weinberger MH (2006) Pathogenesis of salt sensitivity of blood pressure. Curr Hypertens Rep 8:166 –170. [CrossRef] [PubMed] | ||
| 350 | Winnicka K, Bielawski K, Bielawska A, and Miltyk W (2007) Apoptosis-mediated cytotoxicity of ouabain, digoxin and proscillaridin A in the estrogen independent MDA-MB-231 breast cancer cells. Arch Pharm Res 30:1216 –1224. [CrossRef] [PubMed] | ||
| 351 | Xie Z and Askari A (2002) Na+/K+-ATPase as a signal transducer. Eur J Biochem 269:2434 –2439. [CrossRef] [PubMed] | ||
| 352 | Xie Z, Kometiani P, Liu J, Li J, Shapiro JI, and Askari A (1999) Intracellular reactive oxygen species mediate the linkage of Na+/K+-ATPase to hypertrophy and its marker genes in cardiac myocytes. J Biol Chem 274:19323–19328. [CrossRef] [PubMed] | ||
| 353 | Yamada K, Goto A, Nagoshi H, Hui C, and Omata M (1994) Role of brain ouabainlike compound in central nervous system-mediated natriuresis in rats. Hypertension 23:1027–1031. [CrossRef] [PubMed] | ||
| 354 | Yeh BK and Lucchesi BR (1974) Letter: Potassium canrenoate and digitalis intoxication. Circ Res 35:963–967. Yoshika M, Komiyama Y, Konishi M, Akizawa T, Kobayashi T, Date M, Kobatake S, [CrossRef] [PubMed] | ||
| 355 | Masuda M, Masaki H, and Takahashi H (2007) Novel digitalis-like factor, marinobufotoxin, isolated from cultured Y-1 cells, and its hypertensive effect in rats. Hypertension 49:209 –214. [CrossRef] [PubMed] | ||
| 356 | Zahler R, Brines M, Kashgarian M, Benz EJ Jr, and Gilmore-Hebert M (1992) The cardiac conduction system in the rat expresses the α2 and α3 isoforms of the Na+,K+-ATPase. Proc Natl Acad Sci U S A 89:99 –103. Zhang J, Lee MY, Cavalli M, Chen L, Berra-Romani R, Balke CW, Bianchi G, Ferrari [CrossRef] [PubMed] | ||
| 357 | 2005 | P, Hamlyn JM, Iwamoto T, et al | ( 2005 ) Sodium pump alpha2 subunits control myogenic tone and blood pressure in mice. J Physiol 569:243–256. [CrossRef] [PubMed] |
| 358 | Zhang LS, Nakaya K, Yoshida T, and Kuroiwa Y (1991) Bufalin as a potent inducer of differentiation of human myeloid leukemia cells. Biochem Biophys Res Commun 178:686 – 693. [CrossRef] [PubMed] | ||
| 359 | Zouzoulas A, Therien AG, Scanzano R, Deber CM, and Blostein R (2003) Modulation of Na,K-ATPase by the γ subunit: studies with transfected cells and transmembrane mimetic peptides. J Biol Chem 278:40437– 40441. Zvartau NE, Conrady AO, Sviryaev YV, Rotari OP, Merkulova NK, Kalinkin AL, [CrossRef] [PubMed] | ||
| 360 | Shlyakhto EV, and Bagrov AY (2006) Marinobufagenin in hypertensive patients with obstructive sleep apnea. Cell Mol Biol (Noisy-le-grand) 52:24 –27. [CrossRef] [PubMed] | ||
| 361 | Tian J and Xie ZJ (2008) The Na-K-ATPase and calcium-signaling microdomains. Physiology (Bethesda) 23:205–211. Tymiak AA, Norman JA, Bolgar M, DiDonato GC, Lee H, Parker WL, Lo LC, Berova [CrossRef] [PubMed] | ||
| 362 | N, Nakanishi K, and Haber E (1993) Physicochemical characterization of a ouabain isomer isolated from bovine hypothalamus. Proc Natl Acad Sci U S A 90:8189 – 8193. Uddin MN, Horvat D, Glaser SS, Danchuk S, Mitchell BM, Sullivan DE, Morris CA, and Puschett JB (2008a) Marinobufagenin inhibits proliferation and migration of cytotrophoblast and CHO cells. Placenta 29:266 –273. [CrossRef] [PubMed] | ||
| 363 | Uddin MN, Horvat D, Glaser SS, Mitchell BM, and Puschett JB (2008b) Examination of the cellular mechanisms by which marinobufagenin inhibits cytotrophoblast function. J Biol Chem 283:17946 –17953. [CrossRef] [PubMed] | ||
| 364 | Urayama O, Shutt H, and Sweadner KJ (1989) Identification of three isozyme proteins of the catalytic subunit of the Na,K-ATPase in rat brain. J Biol Chem 264:8271– 8280. [CrossRef] [PubMed] | ||
| 365 | Vasilets LA, Postina R, and Kirichenko SN (1999) Mutations of Ser-23 of the α1 subunit of the rat Na+/K+-ATPase to negatively charged amino acid residues mimic the functional effect of PKC-mediated phosphorylation. FEBS Lett 455:8 – 12. [CrossRef] [PubMed] | ||
| 366 | Vasilets LA, Takeda K, Kawamura M, and Schwarz W (1998) Significance of the glutamic acid residues Glu334, Glu959, and Glu960 of the α subunits of Torpedo Na+,K+ pumps for transport activity and ouabain binding. Biochim Biophys Acta 1368:137–149. [CrossRef] [PubMed] | ||
| 367 | Vesely DL, Bayliss JM, and Sallman AL (1987) Human prepro atrial natriuretic factors 26 –55, 56 –92, and 104 –123 increase renal guanylate cyclase activity. Biochem Biophys Res Commun 143:186 –193. Vu H, Ianosi-Irimie M, Danchuk S, Rabon E, Nogawa T, Kamano Y, Pettit GR, Wiese [CrossRef] [PubMed] | ||
| 368 | T, and Puschett JB (2006) Resibufogenin corrects hypertension in a rat model of human preeclampsia. Exp Biol Med (Maywood) 231:215–220. [CrossRef] [PubMed] | ||
| 369 | Vu HV, Ianosi-Irimie MR, Pridjian CA, Whitbred JM, Durst JM, Bagrov AY, Fedorova OV, Pridjian G, and Puschett JB (2005) The involvement of marinobufagenin in a rat model of human peeclampsia. Am J Nephrol 25:520 –528. [CrossRef] [PubMed] | ||
| 370 | Waldorff S and Buch J (1979) Canrenoate—a spironolactone metabolite: acute cardiac effects in digitalized patients. Eur J Cardiol 10:143–149. [CrossRef] [PubMed] | ||
| 371 | Wang H, Haas M, Liang M, Cai T, Tian J, Li S, and Xie Z (2004) Ouabain assembles signaling cascades through the caveolar Na+/K+-ATPase. J Biol Chem 279: 17250 –17259. [CrossRef] [PubMed] | ||
| 372 | Wang H and Leenen FH (2003) Brain sodium channels and central sodium-induced increases in brain ouabain-like compound and blood pressure. J Hypertens 21: 1519 –1524. Wang JG, Staessen JA, Messaggio E, Nawrot T, Fagard R, Hamlyn JM, Bianchi G, and Manunta P (2003) Salt, endogenous ouabain and blood pressure interactions in the general population. J Hypertens 21:1475–1481. [CrossRef] [PubMed] |