Abstract
BACKGROUND Previously we demonstrated that in patients with preeclampsia elevated levels of endogenous Na/K-ATPase inhibitor, marinobufagenin, cause inhibition of Friend leukemia virus integration 1 (Fli1), a negative regulator of collagen-1 synthesis. We hypothesized that in vitro silencing of Fli1 in healthy human umbilical arteries would be associated with an increase in collagen-1 output, similar to the effect of preeclampsia in rat and human tissues.
METHODS The isolated segments of healthy human umbilical arteries were tested for sensitivity to MBG and Fli1 silencing with Fli1 siRNA or control siRNA.
RESULTS Following 24-hour incubation of arteries with nanomolar concentrations of marinobufagenin, Fli1 expression was inhibited 5-fold (P < 0.001), and synthesis of collagen-1 increased 3 times (P < 0.01). Twenty-four-hour incubation of umbilical artery fragments with Fli1 siRNA caused a dramatic decrease of Fli1 (7-fold; P < 0.001) and cytoplasmic PKCδ (4-fold; P < 0.001) expression in comparison to control siRNA or untreated control, followed by elevation in procollagen (3-fold; P < 0.001) and collagen-1 (3-fold; P < 0.001) levels in vascular tissue.
CONCLUSIONS Our results show that after silencing the Fli1 gene in healthy human umbilical arteries a new phenotype emerges which is typical for preeclampsia and is associated with vascular fibrosis.
1. Introduction
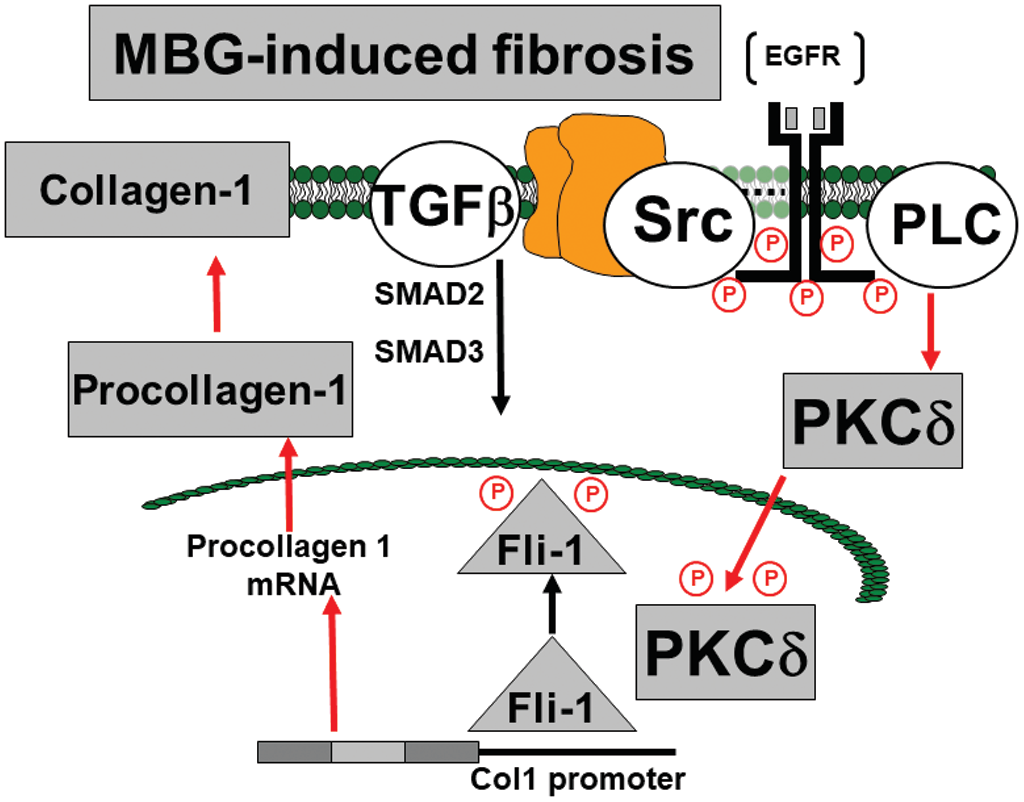
The mechanisms of pathogenesis of preeclampsia (PE) are not well understood, and there is no effective treatment of this disorder.1 Cardiotonic steroid (CS), including marinobufagenin (MBG), a sodium–potassium adenosine triphosphatase (Na/K-ATPase) inhibitor with a high affinity for the alpha-1 isoform of the enzyme, the main isoform in the vascular wall and the kidney, is implicated in PE.2,3 Pregnancy is associated with fluid and sodium retention,4 which is a stimulus for MBG, but MBG levels are not high enough to cause vasospasm.5 In patients with PE, an increase in arterial blood pressure is accompanied by a significant increase in plasma MBG and substantial inhibition of vascular Na/K-ATPase.5,6 The effects of MBG on Na/K-ATPase do not necessarily result in sodium pump inhibition, but rather in the induction of cellular signaling. MBG-induced profibrotic pathway involves Friend leukemia virus integration 1 (Fli1), a nuclear transcription factor that acts as a negative regulator of collagen-1 synthesis.7–9 Fli1 competes with another transcription factor, ETS-1, to maintain a balance between stimulation and repression of the collagen-1 gene.10,11 The Na/K-ATPase/Src/EGF receptor complex begins a signal cascade, which activates phospholipase C (PLC) resulting in the phosphorylation of protein kinase C delta type (PKCδ) and its rapid translocation to the nucleus.11 This translocation is prevented by pharmacological inhibition of PLC.11 In the nucleus, phosphorylated PKCδ phosphorylates Fli1, which withdraws the Fli1-induced inhibition of the collagen-1 promoter and increases procollagen expression and vascular collagen production (Figure 1).11 As a result, PE stiffening of the arterial bed and impairment of vasorelaxation umbilical arteries turns up.9
In recent years, it has become clear that the synthesis of collagen in umbilical arteries represents a part of preeclamptic phenotype12; moreover, tissue fibrosis remains after the recovery from PE.13 Because PE is associated with vascular stiffening,7 we hypothesized that MBG-induced Fli1-dependent synthesis of collagen is implicated in the development of vascular fibrosis and the impairment of vasorelaxation in PE.
Previously, we demonstrated that in human PE level of Fli1 in umbilical arteries is decreased, whereas collagen-1 is elevated, and that ex vivo incubation of preeclamptic arteries with anti-MBG antibody significantly increases Fli1 levels.7 In the present study, we examined (i) whether silencing of the Fli1 gene in healthy human umbilical arteries would produce a preeclamptic phenotype and (ii) whether signaling underlying this process includes Fli1- and transforming growth factor beta (TGF-β)-dependent mechanisms.
2. Methods
General Biological samples (umbilical arteries) and clinical data were collected from subjects with uncomplicated pregnancies in the Department of Obstetrics, School of Pediatric Medicine, St. Petersburg, Russia. The protocol for the human study was approved by the Research Council of St Petersburg School of Pediatric Medicine, Russia, and by the Institutional Review Board of Medstar Research Institute, Washington, DC, renewal on 10 March 2020 (Protocol No. 3/6). All participants read and signed a written informed consent before enrollment. In our study, we enrolled subjects with normotensive, healthy pregnancies (specifically without diabetes mellitus, gestational diabetes). The midsections of the umbilical arteries were immediately placed in ice-cold solution separated from surrounding tissues and divided by rings of 2–3 mm long. Endothelium was removed mechanically, and the rings were rinsed with Dulbecco’s modified Eagle’s medium (DMEM). For MBG treatment, each ring was placed in a separate well of 96-well plate with 250 µl of DMEM supplemented with 10% fetal bovine serum and antibiotics (penicillin, streptomycin, fungizone) and incubated for 48 h at 37 °C and 95% O2/5% CO2. For Fli1 silencing, the solutions of Fli1 siRNA (or scrambled siRNA as a negative control) and transfection reagent were diluted in transfection medium and then mixed with a 1:1 ratio as recommended by the manufacturer. Each umbilical artery ring was incubated in a separate well of a 96-well plate with a final volume of 250 µl of transfection medium. The transfection was conducted with 200 nM Fli1 siRNA (or control siRNA) for 24 h at 37 °C and 95% O2/5% CO2. After 24 hours, the rings were transferred to another plate containing DMEM with 10% fetal bovine serum and antibiotics for a further 24 hours. After incubation, the segments of umbilical arteries were rinsed in DMEM and immediately frozen at −70 °C until use. Frozen tissues were used for the preparation of homogenates. Ten rings incubated in parallel under the same conditions (with the same concentrations of MBG, Fli1 siRNA, or control siRNA) were combined for one sample. Tissues were homogenized in the RIPA buffer lysis system containing protease and phosphatase inhibitors. The homogenates were centrifuged at 11,000 g, and supernatants were collected for immunoblotting. All reagents for transfection and samples processing were purchased from Santa Cruz Biotechnology (USA): Fli1 siRNA (sc-35384), Control siRNA (sc-37007), siRNA Transfection reagent (sc-29528), siRNA Transfection medium (sc-36868), siRNA dilution buffer (sc-29527), and RIPA buffer lysis system (sc-24948).
Western blotting Solubilized proteins were separated by 10–12% Tris–glycine polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane (GE Health Care/Life Sciences, UK). The proteins used included those implicated in profibrotic signaling were visualized using following antibodies: Fli1 (mouse monoclonal, 1:500, Santa Cruz Biotechnology Inc., USA), PKCδ (rabbit polyclonal 1:500, Cell Signaling Technology, USA), phospho-PKCδ (rabbit, 1:500, Cell Signaling Technology, USA), Collagen-1 (goat polyclonal, 1:500, Southern Biotechnology, USA, or mouse monoclonal, 1:500, Santa Cruz Biotechnology, USA), TGF-β (rabbit, 1:500, Cell Signaling Technology, USA), and mothers against decapentaplegic 2 (SMAD-2) (rabbit, 1:500, Sigma–Aldrich, USA). To normalize the levels of proteins against levels of glyceraldehydes-3-phosphate dehydrogenase (GAPDH), the membranes were stripped and reprobed with a mouse monoclonal anti-GAPDH antibody (1:1,000, Santa Cruz Biotechnology, USA) or rabbit polyclonal anti-GAPDH antibody (1:1,000, Cell Signaling Technology, USA), followed by anti-mouse (1:2,000) or anti-rabbit (1:1,000) peroxidase-conjugated antisera (GE Health Care/Life Sciences, UK). Bands were visualized by 1- to 20-minute exposure of nitrocellulose membrane on Premium blue X-ray film (Fenix Research Products, Candler, NC), and optical density was quantified by the laser densitometry (Kodak Molecular Imaging Software, version 5.0, Rochester, NY). MBG was obtained from Bufo marinus toads as previously reported.5
Statistics Data were analyzed using 1-way ANOVA followed by Newman–Keuls test and by 2-tailed t-test (when applicable) (GraphPad Prism software, San Diego, CA). A P-value of less than 0.05 was considered to be statistically significant.
3. Results
Umbilical arteries were collected from 19 nonhypertensive subjects with a mean maternal age of 31 ± 1 years age, 39.0 ± 0.2 weeks gestational age, who underwent delivery via Cesarean section (n = 12) or vaginal delivery (n = 7). The maternal clinical parameters included mean: systolic blood pressure 115 ± 2 mm Hg, diastolic blood pressure 73 ± 1 mm Hg, and a pulse rate of 67 ± 4 beats per minute.
Results of the first experiment in summarized in Figure 2. In this experiment, we studied whether the conditions of treatment of isolated fragments of umbilical arteries, obtained from normotensive women, with nanomolar concentrations of MBG can exert an influence on Fli1 expression and collagen-1 synthesis. As presented in Figure 2, 24-hour incubation of segments of umbilical arteries with nanomolar concentrations of MBG led to dose-dependent decrease in Fli1 expression, and 10 nmol/l MBG significantly increased the level of collagen-1 in umbilical arteries.
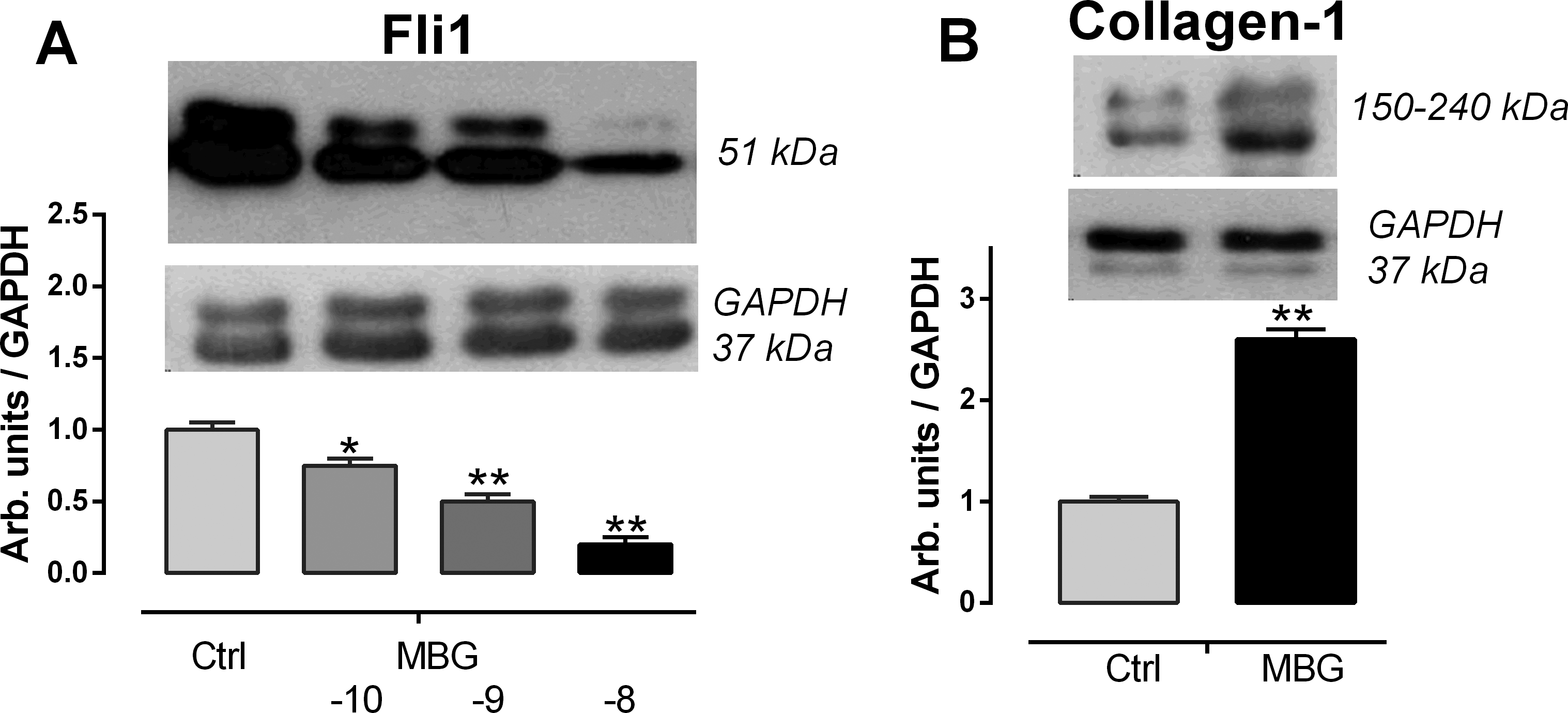
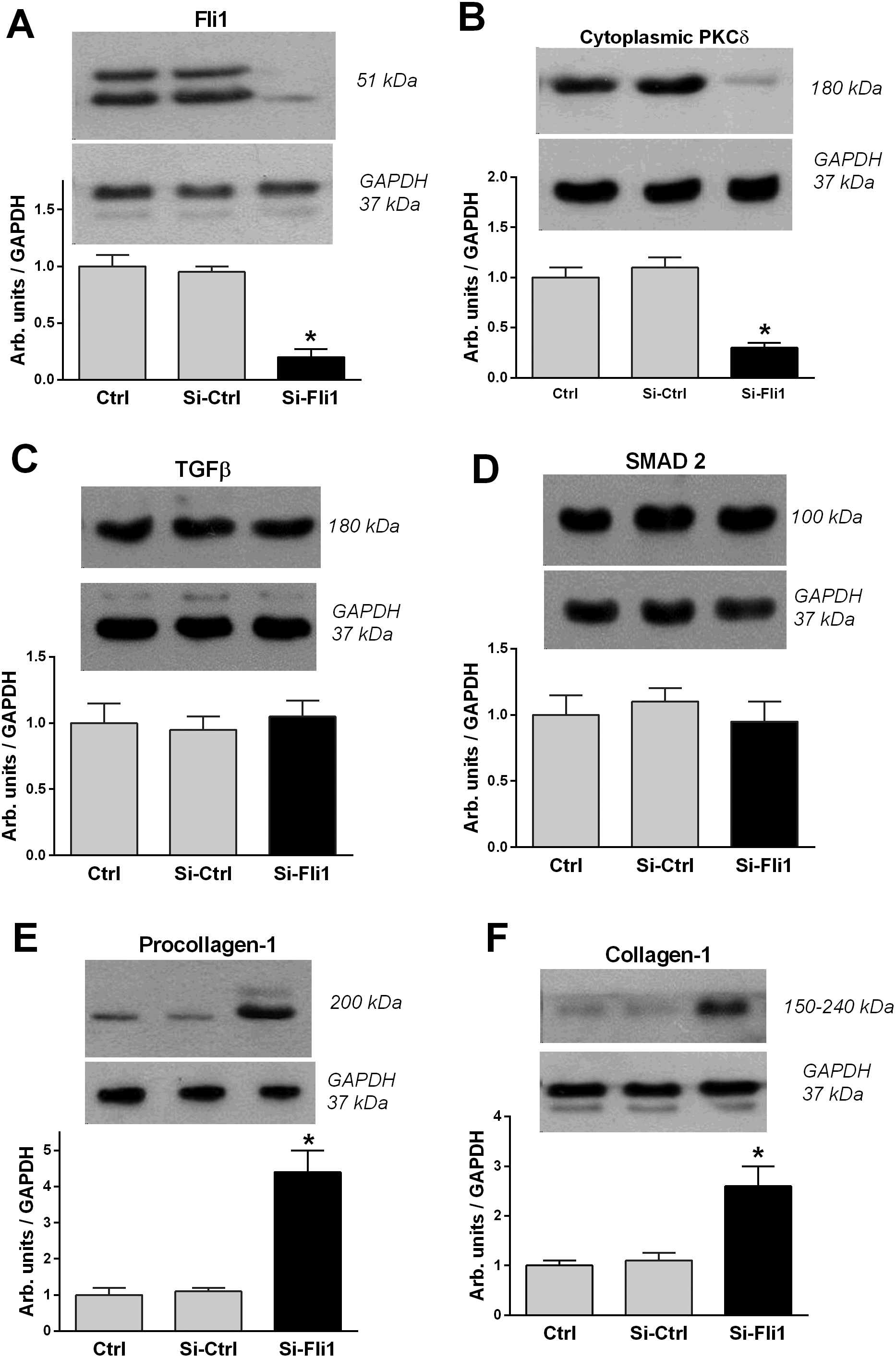
Figure 3 presents the results of experiment in which we assessed the effects of silencing of Fli1 gene. Following silencing of Fli1, its levels reduced 5-fold, which was accompanied by 4-fold reduction of cytoplasmic PKCδ, 3-fold elevation of procollagen-1, and 3-fold increase in collagen-1 content. Control silencing did not change any of the parameters studied. As presented in Figure 3, silencing of Fli1 gene did not produce any effects on levels of TGF-β and SMAD-2, major factors belonging to the transforming growth factor superfamily.
4. Discussion
In the present study, we for the first time showed that silencing of the Fli1 gene in healthy human umbilical arteries results in the emergence of phenotype, which is typical for PE and is associated with the synthesis of collagen. Thus, 24-hour incubation of arterial segments with Fli1 siRNA and low concentrations of MBG resulted in a 5-fold reduction of Fli1 and simultaneous increase in collagen-1. Furthermore, treatment of arterial segments with Fli1 siRNA fully repeated sequence of events observed in PE, dramatic reduction of Fli1, reduction of cytoplasmic PKCδ, and increase in the levels of procollagen and, finally, collagen-1.
PE is a pregnancy disorder with the onset of hypertension after 20 weeks of gestation, with multiple pathophysiologic mechanisms. Several candidates were suggested for the role of endogenous digitalis14,15; of which, MBG has attracted our attention because of several factors. First, in several studies, levels of MBG measured by 4 different antibodies were increased in PE,5,16,17 and second, activity of the Na/K-ATPase in preeclamptic erythrocytes Na/K-ATPase was inhibited and this inhibition was sensitive to addition of anti-MBG antibodies.5,17 Importantly, at concentrations comparable with endogenous levels of MBG in PE, MBG induced vasoconstriction and impaired relaxation of normotensive human umbilical arteries.17 Finally, when pregnant rats were exposed to a high-salt diet, they manifested several symptoms of human PE including fetal growth retardation, proteinuria, hypertension, inhibition of vascular sodium pump, and high plasma renin activity, all of which were reversed by anti-MBG polyclonal and monoclonal antibodies.17,18
The transcription factor, Fli1, a member of the ETS family, is a negative regulator of collagen synthesis, and reduced levels of Fli1 were documented in skin fibroblasts to be also implicated in profibrotic signaling by CS.19 In vitro, we have demonstrated that nanomolar concentrations of MBG stimulate collagen production by dermal, cardiac, and renal fibroblasts by a mechanism involving PKCδ-dependent phosphorylation and depletion of Fli1.11 Low concentrations of CS, acting on PKCδ, induce deactivation of Fli1 and cause expression of collagen-1, which is the key factor of fibrosis in blood vessels, myocardium, and kidney6–8 (Figure 1). Recent studies demonstrate the clinical importance of fibrosis in the cardiovascular system and placenta20,21 and suggest those patients exposed to the PE environment exhibit greater responsiveness to injury despite resolution of PE.21 In addition to MBG-induced Fli1-dependent fibrosis, another profibrotic MBG-produced pathway exists, which is TGF-β and SMAD dependent.7 Notably, in the present study, levels of TGF-β and of SMAD-2 in the umbilical arteries did not change, which indicates that this pathway is not involved in the development of vascular fibrosis in PE and is in accord with the previous findings.22,23
The physiological function of CS is an adaptive response, i.e., regulation of sodium excretion by inhibition of NKA in the epithelial cells of renal tubuli.2 Under pathological conditions, like in patients with PE, the side effect of this primarily adaptive response becomes crucial, and CS causes inhibition of vascular sodium pumps.5,6 Low concentrations of CS acting on PKCδ induce deactivation of Fli1 and cause expression of collagen-1, which is a key factor of fibrosis in blood vessels6,7; however, in the future, effect of silencing of Fli1 should be studied when the force of contractile and relaxant effects of umbilical arteries are measured. In summary, when in healthy umbilical arteries, we reduce levels of Fli1 by siRNA, it mimics PE, and synthesis of collagen-1 becomes dramatically increased.
Funding
The study was supported by Russian Scientific Foundation (grant no. 18-15-00222) and Padakonn Pharma, Narva, Estonia.
Disclosure
The authors declared no conflict of interest.
References
| # | Year | Authors | Title |
|---|---|---|---|
| 1 | 2005 | Sibai B, Dekker G, Kupferminc M | Pre-eclampsia. Lancet 2005; 365:785–799. |
| 2 | 2009 | Bagrov AY, Shapiro JI, Fedorova OV | Endogenous cardiotonic steroids: physiology, pharmacology and novel therapeutic targets. Pharmacol Rev 2009; 61:9–38. |
| 3 | 2018 | Buckalew VM | Role of endogenous digitalis-like factors in the clinical manifestations of severe preeclampsia: a sytematic review. Clin Sci (Lond) 2018;132:1215–1242. |
| 4 | 1994 | Masilamani S, Castro L, Baylis C | Pregnant rats are refractory to the natriuretic actions of atrial natriuretic peptide. Am J Physiol 1994; 267:R1611–R1616. |
| 5 | 1999 | Lopatin DA, Ailamazian EK, Dmitrieva RI, Shpen VM, Fedorova OV, Doris PA, Bagrov AY | Circulating bufodienolide and cardenolide sodium pump inhibitors in preeclampsia. J Hypertens 1999; 17:1179–1187. |
| 6 | 2018 | Fedorova OV, Ishkaraeva VV, Grigorova YN, Reznik VA, Kolodkin NI, Zazerskaya IE, Zernetkina Z, Agalakova NI, Tapilskaya NI, Adair CD, Lakatta EG, Bagrov AY | Antibody to marinobufagenin reverses placenta-induced fibrosis of umbilical arteries in preeclampsia. Int J Mol Sci 2018; 19:2377. |
| 7 | 2020 | Agalakova NI, Reznik VA, Nadei OV, Ershov IA, Rassokha OS, Vasyutina ML, Ivanov DO, Adair ED, Galagudza MM, Bagrov AY | Antibody against Na/K-ATPase inhibitor lowers blood pressure and increases vascular Fli1 in experimental preeclampsia. Am J Hypertens 2020; 33:514–519. |
| 8 | 2012 | Haller ST, Kennedy DJ, Shidyak A, Budny GV, Malhotra D, Fedorova OV, Shapiro JI, Bagrov AY | Monoclonal antibody against marinobufagenin reverses cardiac fibrosis in rats with chronic renal failure. Am J Hypertens 2012; 25:690–696. |
| 9 | 2011 | Nikitina ER, Mikhailov AV, Nikandrova ES, Frolova EV, Fadeev AV, Shman VV, Shilova VY, Tapilskaya NI, Shapiro JI, Fedorova OV, Bagrov AY | In preeclampsia endogenous cardiotonic steroids induce vascular fibrosis and impair relaxation of umbilical arteries. J Hypertens 2011; 29:769–776. |
| 10 | 2007 | Elkareh J, Kennedy DJ, Yashaswi B, Vetteth S, Shidyak A, Kim EG, Smaili S, Periyasamy SM, Hariri IM, Fedorova L, Liu J, Wu L, Kahaleh MB, Xie Z, Malhotra D, Fedorova OV, Kashkin VA, Bagrov AY, Shapiro JI | Marinobufagenin stimulates fibroblast collagen production and causes fibrosis in experimental uremic cardiomyopathy. Hypertension 2007; 49:215–224. |
| 11 | 2009 | Elkareh J, Periyasamy SM, Shidyak A, Vetteth S, Schroeder J, Raju V, Hariri IM, El-Okdi N, Gupta S, Fedorova L, Liu J, Fedorova OV, Kahaleh MB, Xie Z, Malhotra D, Watson DK, Bagrov AY, Shapiro JI | Marinobufagenin induces increases in procollagen expression in a process involving protein kinase C and Fli-1: implications for uremic cardiomyopathy. Am J Physiol Renal Physiol 2009; 296:F1219–F1226. |
| 12 | 2018 | Orabona R, Sciatti E, Prefumo F, Vizzardi E, Bonadei I, Valcamonico A, Metra M, Frusca, T | Pre-eclampsiaheart. a close relationship. Ultrasound Obstet Gynecol 2018; 52:297–301. |
| 13 | 2020 | Melchiorre K, Thilaganathan B, Giorgione V, Ridder A, Memmo A, Khalil A | Hypertensive disorders of pregnancy and future cardiovascular health. Front Cardiovasc Med 2020; 7:59. |
| 14 | 1993 | Graves SW, Glatter KA, Lazarus JM, Williams GH, Hollenberg NK | Volume expansion in renal failure patients: a paradigm for a clinically relevant [Na,K]ATPase inhibitor. J Cardiovasc Pharmacol 1993; 22:S54–S57. |
| 15 | 1994 | Yamada K, Goto A, Hui C, Yagi N, Nagoshi H, Sasabe M, Sugimoto T | Role of ouabainlike compound in rats with reduced renal mass-saline hypertension. Am J Physiol 1994; 266:H1357–H1362. |
| 16 | 2006 | Averina IV, Tapilskaya NI, Reznik VA, Frolova EV, Fedorova OV, Lakatta EG, Bagrov AY | Endogenous Na/K-ATPase inhibitors in patients with preeclampsia. Cell Mol Biol (Noisy-le-grand) 2006; 52:19–23. |
| 17 | 2008 | Fedorova OV, Simbirtsev AS, Kolodkin NI, Kotov AY, Agalakova NI, Kashkin VA, Tapilskaya NI, Bzhelyansky A, Reznik VA, Frolova EV, Nikitina ER, Budny GV, Longo DL, Lakatta EG, Bagrov AY | Monoclonal antibody to an endogenous bufadienolide, marinobufagenin, reverses preeclampsia-induced Na/K-ATPase inhibition and lowers blood pressure in NaCl-sensitive hypertension. J Hypertens 2008; 26:2414–2425. |
| 18 | 2005 | Fedorova OV, Kolodkin NI, Agalakova NI, Namikas AR, Bzhelyansky A, St-Louis J, Lakatta EG, Bagrov AY | Antibody to marinobufagenin lowers blood pressure in pregnant rats on a high NaCl intake. J Hypertens 2005; 23:835–842. |
| 19 | 2010 | Trojanowska M | Cellular and molecular aspects of vascular dysfunction in systemic sclerosis. Nat Rev Rheumatol 2010; 6:453–460. |
| 20 | 2012 | Hausvater A, Giannone T, Sandoval YH, Doonan RJ, Antonopoulos CN, Matsoukis IL, Petridou ET, Daskalopoulou S | The association between preeclampsia and arterial stiffness. J Hypertens 2012; 30:17–33. |
| 21 | 2017 | Orabona R, Sciatti E, Vizzardi E, Bonadei I, Valcamonico A, Metra M, Frusca T | Endothelial dysfunction and vascular stiffness in women with previous pregnancy complicated by early or late pre-eclampsia. Ultrasound Obstet Gynecol 2017; 49:116–123. |
| 22 | 2012 | Bujor AM, Haines P, Padilla C, Christmann RB, Junie M, Sampaio-Barros PD, Lafyatis R, Trojanowska M | Ciprofloxacin has antifibrotic effects in scleroderma fibroblasts via downregulation of Dnmt1 and upregulation of Fli1. Int J Mol Med 2012; 30:1473–1480. |
| 23 | 2022 | Agalakova NI, Grigorova YN, Ershov IA, Reznik VA, Mikhailova EV, Nadei OV, Samuilovskaya L, Romanova LA, Adair CD, Romanova IV, Bagrov AY | Canrenone restores vasorelaxation impaired by marinobufagenin in human preeclampsia. Int J Mol Sci 2022; 23:3336. doi:10.3390/ijms23063336 |