Abstract
Background
Marinobufagenin, NKA (Na/K‐ATPase) inhibitor, causes vasoconstriction and induces fibrosis via inhibition of Fli1 (Friend leukemia integration‐1), a negative regulator of collagen synthesis. In vascular smooth muscle cells (VSMC), ANP (atrial natriuretic peptide), via a cGMP/PKG1 (protein kinase G1)‐dependent mechanism, reduces NKA sensitivity to marinobufagenin. We hypothesized that VSMC from old rats, due to downregulation of ANP/cGMP/PKG‐dependent signaling, would exhibit heightened sensitivity to the profibrotic effect of marinobufagenin.
Methods and Results
Cultured VSMC from the young (3‐month‐old) and old (24‐month‐old) male Sprague–Dawley rats and young VSMC with silenced PKG1 gene were treated with 1 nmol/L ANP, or with 1 nmol/L marinobufagenin, or with a combination of ANP and marinobufagenin. Collagen‐1, Fli1, and PKG1 levels were assessed by Western blotting analyses. Vascular PKG1 and Fli1 levels in the old rats were reduced compared with their young counterparts. ANP prevented inhibition of vascular NKA by marinobufagenin in young VSMC but not in old VSMC. In VSMC from the young rats, marinobufagenin induced downregulation of Fli1 and an increase in collagen‐1 level, whereas ANP blocked this effect. Silencing of the PKG1 gene in young VSMC resulted in a reduction in levels of PKG1 and Fli1; marinobufagenin additionally reduced Fli1 and increased collagen‐1 level, and ANP failed to oppose these marinobufagenin effects, similar to VSMC from the old rats with the age‐associated reduction in PKG1.
Conclusions
Age‐associated reduction in vascular PKG1 and the resultant decline in cGMP signaling lead to the loss of the ability of ANP to oppose marinobufagenin‐induced inhibition of NKA and fibrosis development. Silencing of the PKG1 gene mimicked these effects of aging.
Keywords: aging, atrial natriuretic peptide, Na‐pump inhibitor marinobufagenin, PKG1 gene silencing, vascular smooth muscle cells
Nonstandard Abbreviations and Acronyms
| FBS | fetal bovine serum |
| Fli1 | Friend leukemia integration‐1 |
| NKA | Na/K‐ATPase |
| PKG1 | protein kinase G1 |
| VSMC | vascular smooth muscle cells |
Clinical Perspective
What Is New?
Age‐associated reduction in vascular PKG1 (protein kinase G1) levels and resultant decline in cGMP‐dependent signaling sensitize vascular smooth muscle cells to the profibrotic effect of marinobufagenin.Silencing of the PKG1 gene in vascular smooth muscle cells from young rats mimics these effects of aging.
What Are the Clinical Implications?
The observed shift between vascular and renal effects of marinobufagenin underlies an age‐dependent increase in the salt‐sensitivity of blood pressure.
Salt sensitivity contributes to elevated blood pressure (BP) in 30% to 40% of people with hypertension worldwide.1, 2 Endogenous cardiotonic steroids, like marinobufagenin, are stimulated by renal sodium retention and are implicated in the response to NaCl loading.3, 4 Alpha‐1 NKA (Na/K‐ATPase) isoform exhibits heightened marinobufagenin sensitivity and is an exclusive NKA isoform in the renal tubules and major isoform in vascular smooth muscle cells (VSMC).5, 6 In people with normal BP, marinobufagenin, acting via inhibition of the renal Na pump, promotes sodium excretion; but in subjects who are salt‐sensitive the blunted renal Na pump function is one of the factors underlying renal Na retention.4, 7 In these subjects, excessive production of marinobufagenin occurs to override sodium retention, but marinobufagenin exhibits a maladaptive effect, inhibits NKA in the vascular sarcolemma, and raises BP.7, 8 Previously it was demonstrated that plasma levels of marinobufagenin in humans and in rodent models range from 0.1 to 2 nmol/L, that is, physiological marinobufagenin level, and it is higher in preeclampsia, chronic kidney disease, and salt‐sensitive hypertension.7, 8, 9, 10, 11, 12
Previously, we demonstrated that in the VSMC, ANP (atrial natriuretic peptide), acting via a cGMP/PKG1 (protein kinase G1)‐dependent mechanism, reduces the sensitivity of NKA to marinobufagenin.13 PKG1 is a serine/threonine‐specific protein kinase that is activated by cGMP. We also found that compared with 3‐month‐old rats, NaCl‐loaded adult (12‐month‐old) rats exhibited a greater marinobufagenin response, greater BP elevation, greater inhibition of NKA in the aorta, and 30% less natriuresis in the presence of comparable responses of α‐hANP (alpha‐human atrial natriuretic polypeptide) and cGMP.13 In that study, the levels of PKG were lower, whereas the levels of phosphodiesterase‐5 were higher in the aorta and kidneys of older compared with young rats, and α‐hANP failed to modulate marinobufagenin‐induced NKA inhibition in older rats.13 These results suggest that age‐associated downregulation of cGMP/PKG‐dependent signaling impairs the ability of ANP to modulate the effects of marinobufagenin on the sodium pump, which contributes to salt‐sensitivity of BP.
The contribution of marinobufagenin to the pathogenesis of hypertension, however, is not limited to its vasoconstrictor effects, because a growing body of evidence indicates that physiological, or nanomolar, concentrations of this steroidal NKA inhibitor stimulate the synthesis of collagen‐1 and induce cardiac, vascular, and renal fibrosis via binding to NKA and initiation of the downstream profibrotic signaling.14, 15, 16 One of these pathways is marinobufagenin‐induced fibrosis, which occurs via PKCσ (protein kinase C delta)‐dependent inhibition of Fli1 (Friend leukemia integration‐1), a nuclear transcription factor and a negative regulator of collagen synthesis.4, 17
Because fibrosis is a hallmark of cardiovascular aging, and because aging is associated with downregulation of cGMP/PKG‐dependent mechanisms, we hypothesized that (1) VSMC from old rats would exhibit heightened sensitivity to the profibrotic effect of marinobufagenin, and (2) silencing of PKG1 gene in VSMC from young rats would mimic this effect.
METHODS
The data that support the findings of this study are available from the corresponding author upon reasonable request.
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Animal Care and Use Committee of National Institute on Aging, National Institutes of Health (protocol code 191‐LCS, approved January 11, 2012).
Experimental Model
The experimental protocol was approved by the Animal Care and Use Committee of the National Institute on Aging. Three‐month‐old (n=32) and 24‐month‐old (n=32) male Sprague–Dawley rats (Charles River Laboratories, Wilmington, MA) were used in the experiments. Eight rats from each age group were used for VSMC preparation. The other rats (3‐month‐old; n=24; 321±7 g and 24‐month‐old; n=24; 560±16 g) were studied after 1 week of adaptation to laboratory environment, metabolic chambers, and measurement of blood pressure via tail‐cuff plethysmography (IITC Inc., Life Science, Woodland Hills, CA). Following baseline urine collection and blood pressure measurement 12 young and 12 old rats were euthanized, serving as controls, and a single injection of 2.5 mL/kg 20% NaCl was intraperitoneally administered to 12 rats of each age group under light anesthesia (25 mg/kg Ketaject).13 The animals were placed in the metabolic chambers, and urine was collected hourly for estimation of renal excretion of sodium, marinobufagenin and cGMP. Within 2 hours following acute NaCl‐loading, systolic blood pressure (SBP) was measured, and the animals were anesthetized with 60 mg/kg Ketaject and exsanguinated from the abdominal aorta. Plasma was collected for determination of marinobufagenin and α‐ANP‐1‐28. Concentration of sodium in the urine was measured using Roche‐Hitachi 917 (Roche, Vienna, Austria), and urinary creatinine was measured by a colorimetric method using a creatinine urinary assay kit (Cayman Chemical, Ann Arbor, MI). Plasma electrolytes, plasma creatinine, and plasma glucose were measured using commercially available i‐STAT cartridges Crea and CHEM 8+ (Abbott Point of Care, NJ).
The aortae and kidneys were fixed in 4% buffered formalin and embedded in paraffin. After the removal of paraffin and rehydration, aortic sections (6 μm thick) were stained with Verhoeff Elastin Staining Kit (American Mastertech, Lodi, CA) and kidney sections (6 μm thick) were attained with Masson’s Trichrome stain (American Mastertech). The slides were then dehydrated 3 times in 100% ethanol, cleared in xylene, and mounted with Permount (Fisher Scientific, Asheville, NC). The photomicrographs were taken by Zeiss Axioplan microscope and Qicam digital camera (Thornwood, NY).
Vascular Smooth Muscle Cells Preparation
The aortae from 3‐month‐old (n=8) and 24‐month‐old (n=8) control Sprague–Dawley rats were used for VMSC preparation as described18, 19 with minor modifications. Briefly, the aortae were incubated with collagenase type I (Worthington Biochemical Corporation, Lakewood, NJ) in Hank’s balanced salt solution (Life Technologies, Grand Island, NY) with antibiotic mixture (penicillin and streptomycin with an addition of antibiotic‐antimycotic mixture; Life Technologies) at 37 °C for 30 minutes. After the removal of adventitia and endothelial cells, the tissues were incubated in DMEM (Life Technologies) with 10% fetal bovine serum (FBS; Life Technologies), and antibiotic mixture at 37 °C overnight. Next, the tissues were minced with scissors and incubated for 1.5 hours with elastase (0.5 mg/mL; Sigma, St. Louis, MO) and collagenase type II (Worthington Biochemical Corporation) in Hank’s balanced salt solution buffer with antibiotics. Digestion was stopped by adding DMEM with 10% FBS and antibiotics. The cells were filtered through a 70 μm cells strainer, collected by centrifugation (500g for 5 minutes), and plated on collagen‐covered tissue culture plate in DMEM with 10% FBS with antibiotics as described20 with minor modifications.19 The young and old VSMC were passaged 4 times until the cells reached the number high enough to perform the treatment. VSMC from 3‐month‐old rats (fourth passage; ~90% confluence19) were used for silencing of PKG1 gene. Both VSMC from 3‐month‐old (young cells) and 24‐month‐old rats (old cells) from fourth passage were plated at the same density ~90% confluence and were depleted in FBS (1%) for 24 hours followed by 24 hours incubation with 1 nmol/L marinobufagenin or α‐hANP 1 nmol/L or combination of both compounds. After 24 hours, the VSMC were used for Western blotting analysis.
Western Blotting
Solubilized proteins from aortic sarcolemma were separated by 10% Tris‐glycine polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane (GE Healthcare/Life Sciences). The proteins were visualized using polyclonal rabbit anti‐PKG1 (cGPK1β) antibody (Stressgen, Victoria, BC, Canada, 1:2000), rabbit polyclonal anti‐Fli1 antibody (1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), goat polyclonal anti‐collagen‐1 antibody (1:500; Southern Biotechnology, Birmingham, AL) followed by incubation with peroxidase‐conjugated anti‐mouse, anti‐rabbit antiserum (Amersham Corp., 1:1000), or donkey anti‐goat antiserum (Santa Cruz Biotechnology, Inc., 1:1000). Bands were visualized by 1‐ to 30‐minute exposure of nitrocellulose membrane to Premium Blue X‐ray film (Phenix Research Products at Thomas Scientific, Swedesboro, NJ), and optical density was estimated by laser densitometry (Kodak Molecular Imaging Software, version 5.0). To normalize levels of proteins against levels of GAPDH or α‐tubulin, membranes were stripped and reprobed with rabbit monoclonal anti‐GAPDH antibody (Cell Signaling Technology, Danvers, MA; 1:1000) or with monoclonal mouse anti‐α‐tubulin (Sigma‐Aldrich, St. Louis, MO; 1:5000).
Immunoassays
Plasma and urine samples were extracted on Sep‐Pak C‐18 cartridges (Waters, Milford, MA), and marinobufagenin competitive fluoroimmunoassay based on a monoclonal murine anti‐marinobufagenin 4G4 antibody was performed as described recently in detail.13 The cross‐reactivity of 4G4 anti‐marinobufagenin antibody is as follows: marinobufagenin—100%, ouabain—0.005%, digoxin—0.03%, digitoxin <0.001%, bufalin—0.08%, cinobufagin—0.07%, cinobufatalin—40%, prednisone <0.001%, spironolactone <0.001%, proscillaridin <0.001%, and progesterone <0.001%. Plasma levels of rat ANP 1–28 were measured using enzyme immunoassay kit (Peninsula Laboratories, San Carlos, CA). Urinary cGMP was measured using enzyme immunoassay kit (Cayman Chemical, Ann Arbor, MI).
Silencing of PKG1 Gene
VSMC from young (3 months old) rats were used for silencing of PKG1 gene. Double‐stranded siRNAs were used for the silencing of PRKG1 (L‐085799‐01‐0050; former Dharmacon Inc., present Horizon Discovery Ltd.); ON‐TARGETplus Non‐targeting Pool (D‐001810‐10‐20; Dharmacon Inc.) was used as a negative control. The transfection of VSMC was conducted with 100 nmol/L siRNA using Gene Silencer as a transfection reagent (T500750; Genlantis Inc., San Diego, CA) according to the manufacturer’s instructions. Transfection was made during 6 hours in DMEM; after 6 hours incubation FBS was added to the final concentration 10%; medium was completely changed after 24 hours of incubation with siRNA to DMEM with 10% FBS and antibiotics. Cells were collected for the Western blot analysis after the inclubation with 1 nmol/L marinobufagenin or α‐hANP 1 nmol/L or combination of both compounds following 96 hours after transfection with si‐PKG1.
Na/K‐ATPase Activity
NKA activity was measured as reported previously.13 To increase the permeability of membrane vesicles, membranes were pretreated with alamethicin (0.5 mg/mg protein). Aliquots of membranes from aortic sarcolemma (0.5 μg protein/100 μL) were preincubated for 60 minutes at 37 °C with marinobufagenin (1 nmol/L) as described and then incubated for 1 hour in the absence and presence of α‐hANP 1 nmol/L at 37 °C in NUNC low binding polystyrene plates in medium containing (in mmol/L): NaCl 100; KCl 4; MgCl2 3; EDTA 1; Tris 50; ATP 3; and NaN3 5; pH 7.4. The reaction was stopped by adding the quenching solution containing 1.25 N H2SO4. Total ATPase activity was measured by the production of inorganic phosphate, and NKA activity was estimated as a difference between ATPase activity in the presence and in the absence of 5 mmol/L ouabain.
Statistical Analysis
Results are reported as the mean±SEM. Shapiro–Wilk normality tests (GraphPad Prism software; GraphPad Inc, San Diego, CA) were conducted for each sample and each variable. All samples passed the normality test (α>0.05). Because our data were consistent with normal distributions with constant variance, the significance of differences among the measured variables was assessed by 1‐way ANOVA followed by Newman–Keuls test, 2‐way ANOVA followed by Holm‐Sidak’s multiple comparisons test, or t test, where applicable (GraphPad Prism software; GraphPad Inc.). A 2‐sided P value of <0.05 was considered significant.
RESULTS
Data on the levels of plasma marinobufagenin and ANP and vascular levels of PKG1 and Fli1 in young and old rats are summarized in Figure 1. Twenty‐four‐month‐old animals had similar plasma marinobufagenin levels and lower plasma ANP levels, lower aortic PKG1, and lower Fli1 compared with 3‐month‐old rats (Figure 1). The Table lists the baseline levels of urine excretion, plasma electrolytes, hematocrit, and plasma glucose; these parameters did not differ between young and old rats.
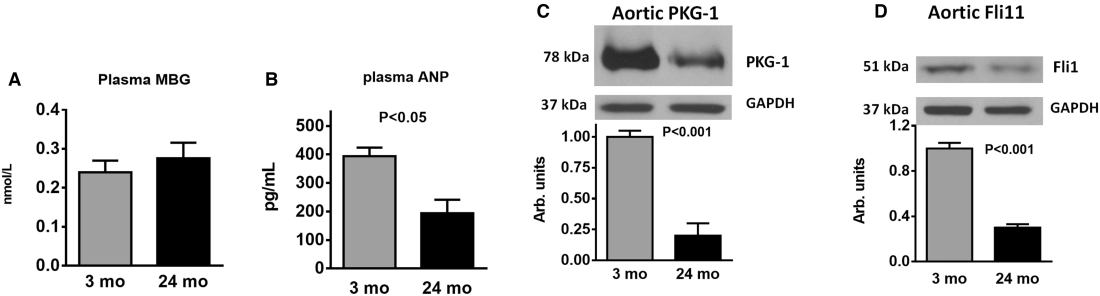
Table 1
Blood Pressure, Urine Excretion, Plasma Electrolytes, and Hematocrit in Young and Old Sprague–Dawley Rats
| 3‐month‐old | 24‐month‐old | |
|---|---|---|
| Systolic blood pressure, mm Hg | 125±3 | 127±4 |
| Urine excretion, mL/h | 5.8±0.7 | 4.6±0.76 |
| Plasma Na, mmol/L | 143±0.5 | 144±0.6 |
| Plasma K, mmol/L | 4.2±0.1 | 4.1±0.1 |
| Hematocrit, % | 41±0.5 | 42±0.7 |
| Plasma glucose, mg/dL | 13.8±0.2 | 14.1±0.2 |
Table 1
Blood Pressure, Urine Excretion, Plasma Electrolytes, and Hematocrit in Young and Old Sprague–Dawley Rats
| 3‐month‐old | 24‐month‐old | |
|---|---|---|
| Systolic blood pressure, mm Hg | 125±3 | 127±4 |
| Urine excretion, mL/h | 5.8±0.7 | 4.6±0.76 |
| Plasma Na, mmol/L | 143±0.5 | 144±0.6 |
| Plasma K, mmol/L | 4.2±0.1 | 4.1±0.1 |
| Hematocrit, % | 41±0.5 | 42±0.7 |
| Plasma glucose, mg/dL | 13.8±0.2 | 14.1±0.2 |
Representative images of the histological analyses of aorta and kidney from young and old rats are presented in Figure 2. The cross section of aorta from the 3‐month‐old rats revealed moderate collagen label (purple color) with Verhoeff’s staining in the area of the external elastic lamina and the adventitia layer (Figure 2A). The medial part of the aortic wall of the 24‐month‐old rats was thicker, the VSMC were hypertrophied, and the accumulation of collagen was obvious and more pronounced in the media layer between the smooth muscle cells compared with the aorta from the young rats (Figure 2B).
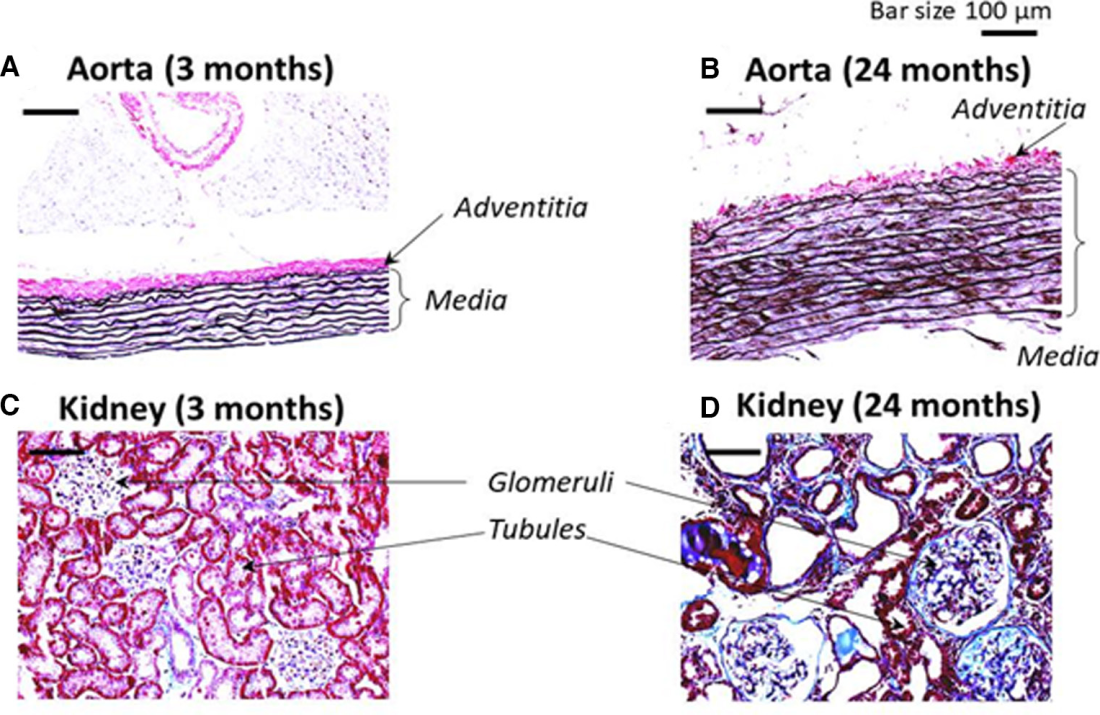
Masson’s trichrome stain of kidney from the 3‐month‐old rats demonstrated that most renal tubules and the basement membrane of Bowman’s capsule contain a thin layer of collagen (Figure 2C). The renal vessels displayed collagen staining mainly in the adventitia. In the kidney of the 24‐month‐old rat collagen was accumulated markedly in the wall of small and large vessels, and the kidney was hypertrophied with degenerated renal tubules. Renal tubules have thickened walls and most of the glomeruli in the corpuscles were shrunken in 24‐month‐old kidney (Figure 2D). These changes in collagen abundance in both aorta and kidneys indicate that the old rats developed fibrosis in these tissues in comparison to the young animals.
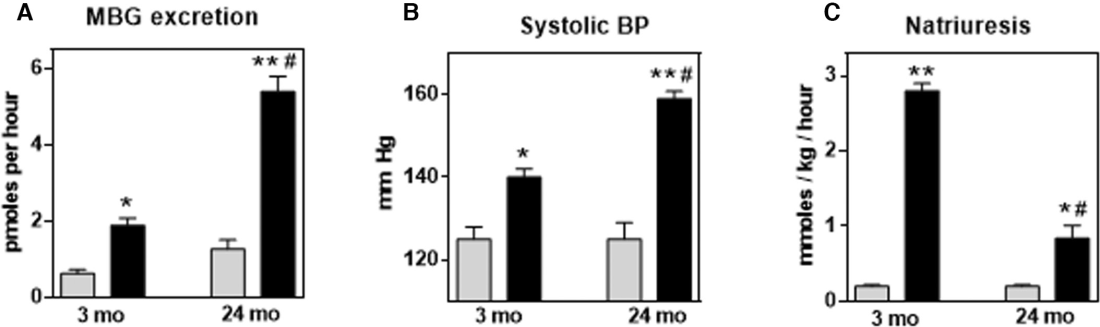
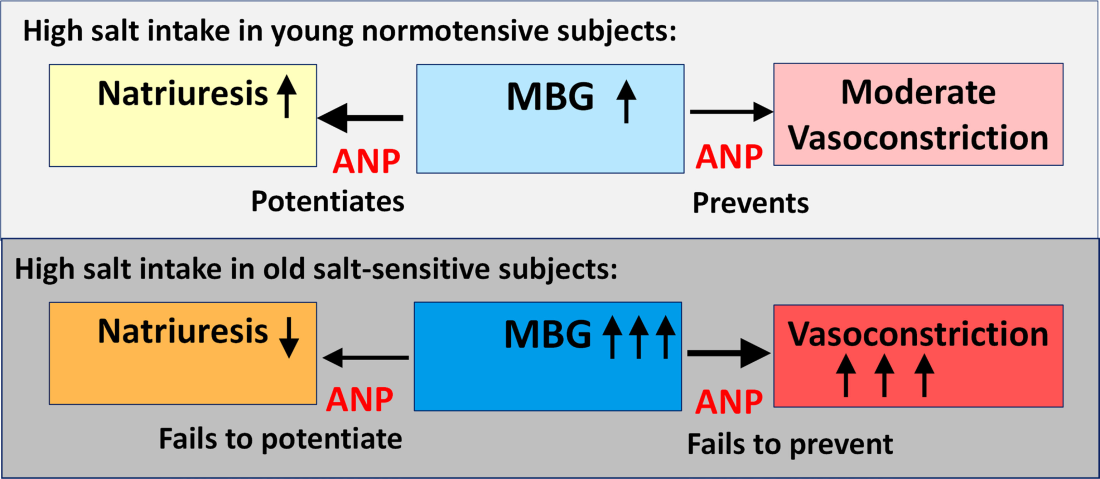
Acute NaCl loading was accompanied by an increase in urine marinobufagenin excretion, SBP, and natriuresis in both young and old rats (Figure 3). Twenty‐four‐month‐old rats exhibited greater marinobufagenin and pressor responses (Figure 3A and 3B) and a blunted natriuretic response (Figure 3C) after acute NaCl loading compared with 3‐month‐old rats, indicating that the age‐associated changes in the old rats, that is, downregulation of PKG1 and Fli1 (Figure 1), and the development of aortic and renal fibrosis (Figure 2) may underlie the age‐associated increase in salt sensitivity.
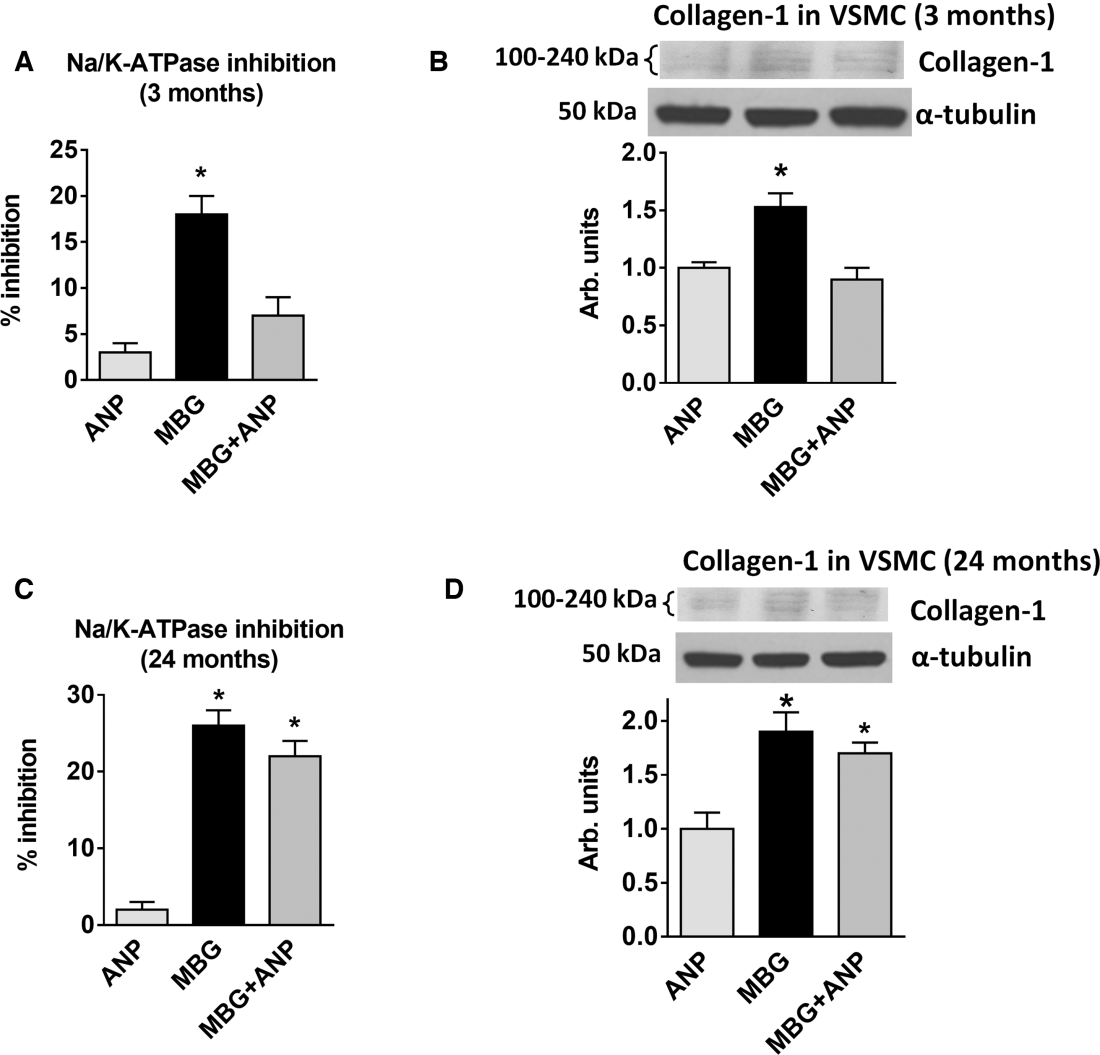
We previously demonstrated that the rats with NaCl‐induced hypertension had plasma marinobufagenin concentration in the nanomolar range.7 Figure 4 illustrates the modulatory effects of a 24‐hour administration of α‐hANP (1 nmol/L) on NKA inhibition by 1 nmol/L marinobufagenin on collagen‐1 synthesis in VSMC from young and old rats. In 3‐month‐old rats, 1 nmol/L α‐hANP markedly reversed marinobufagenin‐induced NKA inhibition in VSMC and prevented an increase in vascular levels of collagen‐1, even in the presence of marinobufagenin (Figure 4A and 4B). However, in VSMC from the old rats, α‐hANP failed to modulate NKA‐inhibitory and collagen‐1 stimulatory effect of marinobufagenin (Figure 4C and 4D). Thus, in the older rats, in vitro, 1 nmol/L α‐hANP failed to reduce the effect of marinobufagenin on NKA in VSMC and did not reduce marinobufagenin‐induced increase of collagen‐1, as it did in the young rats.
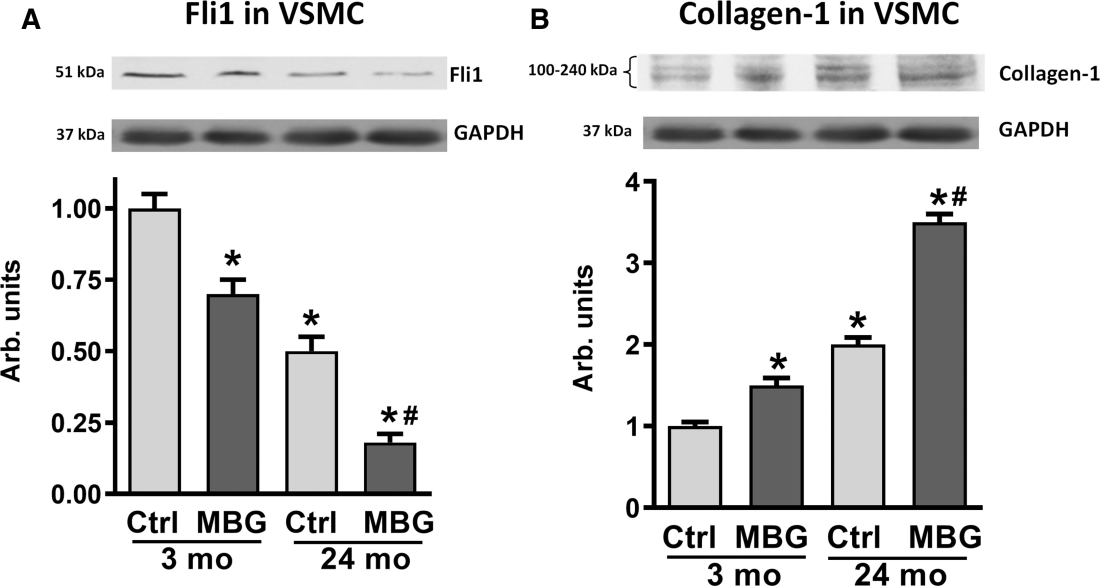
Next we investigated the involvement of a negative regulator of collagen production, Fli1, in the age‐dependent change of vascular levels of collagen‐1. Figure 5 shows, that whereas in 3‐month‐old rats, 1 nmol/L marinobufagenin induced inhibition of Fli1 and concurrent stimulation of collagen‐1 in the aortic VSMC versus control nontreated cells, the magnitudes of Fli1 and collagen‐1 changes were greater in VSMC from the 24‐month‐old rats (Figure 5). Notably, that Fli1 was lower and collagen‐1 was higher in control 24‐month‐old VSMC versus 3‐month‐old VSMC.
We next determined (1) whether a reduction in the level of expression of PKG1 via gene silencing in VSMC from young rats would mimic the age‐associated changes in sensitivity of vascular NKA to marinobufagenin; and (2) whether the reduction of PKG1 expression in the young VSMC would promote the profibrotic effect of marinobufagenin on Fli1 and collagen‐1, observed in the old rats (Figure 6).
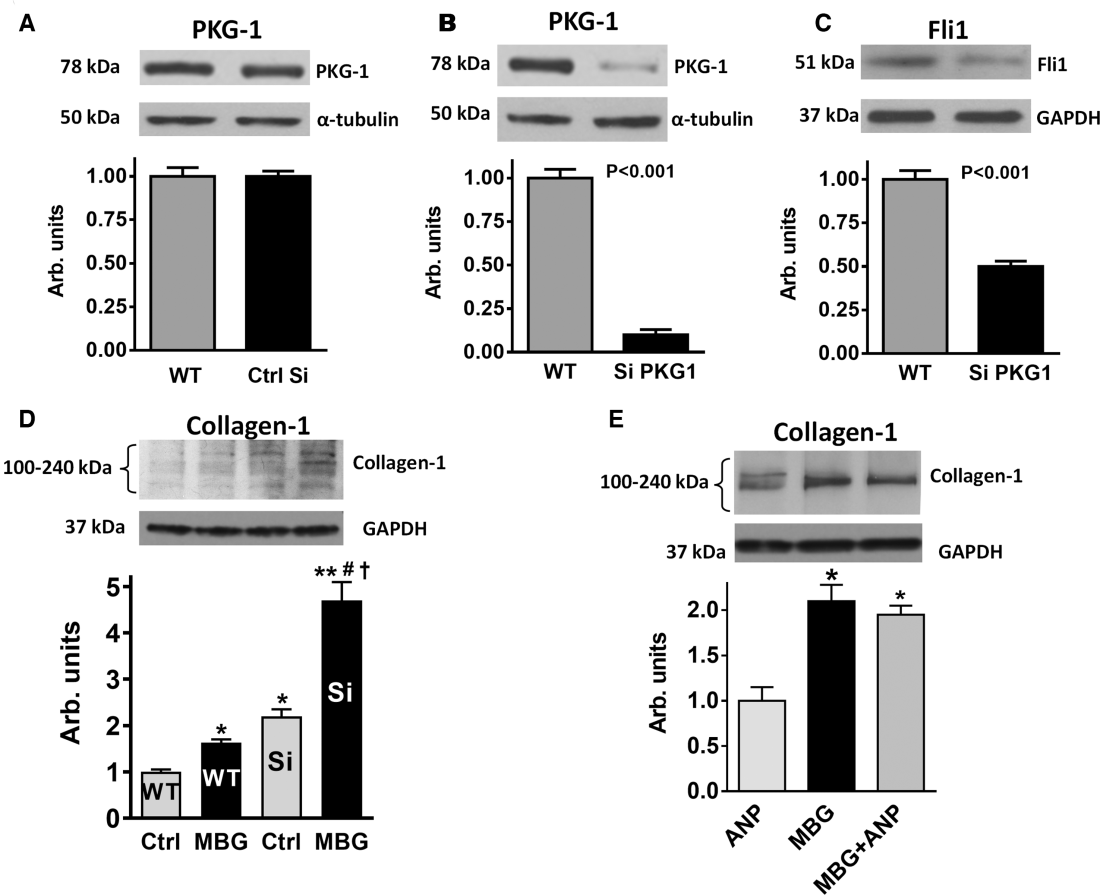
Although nontarget (control) silencing did not affect the expression of PKG1 in the aortic VSMC cells (Figure 6A), silencing of PKG1 by siRNA dramatically reduced level of PKG1 immunoreactivity (Figure 6B). Interestingly, the levels of Fli1 also decreased (Figure 6C) and levels of collagen‐1 in VSMC increased following PKG1 gene silencing (Figure 6D, wild type control vs silencing of PKG1 gene [siPKG1] control). Next we compared the effect of marinobufagenin on the synthesis of collagen‐1 in control VSMC and in VSMC following the silencing of PKG1 (Figure 6D); the incubation of control VSMC with 1 nmol/L marinobufagenin led to the moderate increase in collagen‐1 level; silencing of PKG1 was associated with an increase in collagen‐1, and treatment with 1 nmol/L marinobufagenin further increased collagen‐1 production in VSMC with silenced PKG1 gene. α‐hANP failed to modulate the effect of marinobufagenin in the cells with silenced PKG1 gene (Figure 6E).
DISCUSSION
Progression of fibrosis, downregulation of cGMP‐dependent protective mechanisms, and development of salt‐sensitive hypertension are the hallmarks of cardiovascular aging; we studied the role of marinobufagenin and PKG1 in vascular aging. The main findings of the present study are (1) the development of age‐associated salt sensitivity in normotensive rats is accompanied by an exaggerated response of marinobufagenin to NaCl loading due to a marked reduction levels of PKG1 and Fli1 in the vasculature; (2) VSMC from 24‐month‐old rats (versus 3‐month‐old animals) exhibit an increased sensitivity to the profibrotic effect of marinobufagenin; and (3) silencing of PKG1 gene in VSMC from young rats mimics this effect, that is, silencing of PKG1 gene makes VSMC prone to the profibrotic action of marinobufagenin, whereas ANP loses its ability to protect the cells from that effect of marinobufagenin. This finding indicates that PKG1 plays an important role in modulation of profibrotic effect of marinobufagenin via activation of an ANP‐dependent protective mechanism.
We previously demonstrated that the impact of effects of ANP on marinobufagenin‐induced NKA inhibition in aortic sarcolemma from aged male Sprague–Dawley rats resembles that in the Dahl rats.7 Whereas in young Sprague–Dawley rats salt loading did not elevate SBP and ANP reversed the inhibition of vascular NKA, this effect was absent in aged rats, in which ANP failed to offset marinobufagenin‐induced NKA inhibition in the aortic sarcolemma (Figure 7), similar to Dahl rats, in which a pressor effect was associated with a lesser response of cGMP and ANP and significantly reduced renal sodium excretion.7 Moreover, silencing of PKG1 in VSMC from 3‐month‐old rats converted the “young phenotype” to an “old phenotype,” characterized by down‐regulation of PKG1 gene and the resultant reduction in cGMP‐dependent signaling, leading to the loss of the ability of ANP to modulate interactions between marinobufagenin and NKA. As previously demonstrated in the older rats, ANP fails to reduce marinobufagenin‐induced inhibition of the NKA in the vasculature leading to vasoconstriction.15 Several studies in humans have shown positive correlations between pulse wave velocity, an index of arterial stiffness, and urinary marinobufagenin in subjects of both sexes with moderately increased BP and pulse wave velocity,21 in high and low salt intake in male patients,9 and the positive relationship between pulse wave velocity and ratio between marinobufagenin‐dependent erythrocyte NKA activity during normal Na and salt loading in male patients with resistant hypertension.8 Interestingly, no such relationship between pulse wave velocity and marinobufagenin was detected in the 2 last studies in female subjects.8, 9
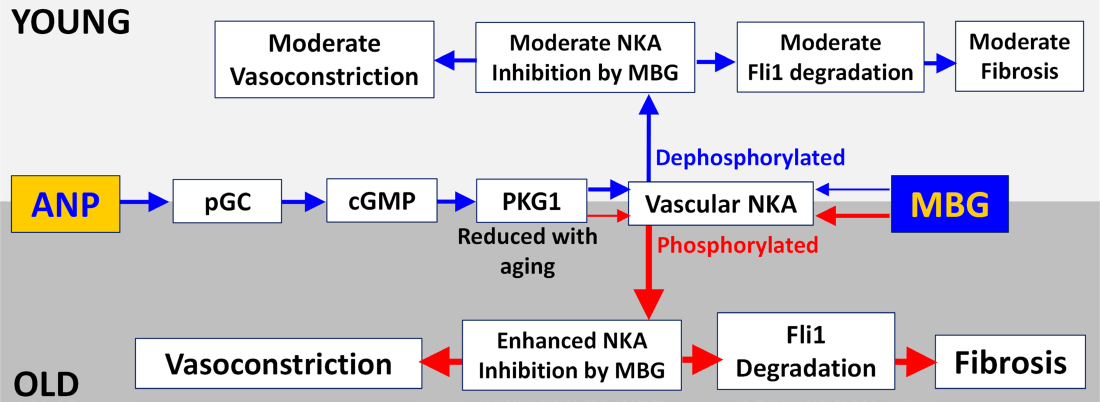
In addition to a greater sensitivity of their arteries to the contractile effect of marinobufagenin, old rats are becoming sensitive to the profibrotic effect of marinobufagenin and even lower concentrations of marinobufagenin were able to induce fibrosis in the aged cells. This is based on the ability of marinobufagenin to inhibit NKA and to initiate tissue remodeling via the profibrotic Fli1‐dependent signaling.4
Fli1 is the transcription factor and a negative regulator of collagen production, which is involved in marinobufagenin‐induced fibrosis.14 Following inhibition of SRC (proto‐oncogene tyrosine‐protein kinase) and phospholipase C by marinobufagenin, activation of PKCδ phosphorylates Fli1 leading to procollagen‐induced synthesis.22, 23 Thus, mice with 5/6 partial nephrectomy significantly increased left ventricular fibrosis.23 Notably, although in previous studies this chain of events has been demonstrated to occur in both cardiac or renal fibroblasts,23 the present results show that it also occurs in the VSMC.15 Because 2 signaling pathways are known to modulate the profibrotic effects of marinobufagenin, a question arises whether the profibrotic effect of marinobufagenin may be initiated via the SMAD (mothers against decapentaplegic homolog)‐dependent TGFβ‐1 (transforming growth factor beta‐1) signaling24 in addition to the Fli1‐dependent mechanism.
The development of age‐associated salt sensitivity of BP in Sprague–Dawley rats is accompanied by an increased sensitivity of vascular sodium pump to marinobufagenin, down‐regulation of PKG1 in the vasculature, and a resultant loss of the ability of ANP to reduce marinobufagenin‐induced inhibition of NKA appears to underlie this effect (Figure 7). In VSMC cGMP binds to pathway effector molecules, including PKG1 protein and phosphodiesterases.25, 26 These mediators participate in a variety of physiological actions in the context of hypertension, including SBP increase, increased in glomerular filtration rate and natriuresis, and collagen‐1 upregulation, thus opposing vascular cellular injury and remodeling.25, 26 cGMP, produced by guanylate cyclase, activates PKG1 and regulates vascular remodeling, schematically presented in Figure 7.
Thus, our present findings are in agreement with the growing body of evidence that, concerning the pathogenesis of hypertension, the effects of endogeonous steroidal NKA inhibitors on growth and collagen synthesis (vascular stiffness) may be more important than their effects on SBP. Analysis of our recent human data suggests that the involvement of marinobufagenin in response to a NaCl challenge becomes altered with advancing age and that there is a sex difference in the response of cardiac steroids to salt loading.9, 27 Whereas in older women, a relative failure in marinobufagenin production may cause volume expansion and contribute to hypertension, in men, aging is associated with lower baseline marinobufagenin levels than in women but a greater responsiveness of marinobufagenin to the administration of a high salt diet.9, 21
CONCLUSIONS
In summary, in VSMC from young and old rats, preincubation with marinobufagenin was associated with an increased level of collagen‐1, and this effect of marinobufagenin became stronger in advanced age, that is, the older VSMC exhibited higher collagen level after marinobufagenin administration than did young VSMC. The observed age‐associated effect of marinobufagenin was ANP/PKG‐dependent, because silencing of the PKG1 gene in young VSMC was associated with augmentation of the marinobufagenin effect of collagen‐1 level. ANP failed to protect VSMC against the profibrotic effect of marinobufagenin following silencing of the PKG1 gene. Thus, the ANP/PKG‐dependent mechanism of augmentation of the effects of marinobufagenin very likely contributed to the development of salt sensitivity of BP, accompanied by the development of marinobufagenin‐induced cardiovascular fibrosis. Accordingly, following acute NaCl loading, 24‐month‐old rats exhibited a greater pressor response and a lesser natriuretic response to NaCl loading as compared with 3‐month‐old rats (Figure 8; schematic presentation of marinobufagenin implication in age‐associated salt sensitivity). Our present data are in agreement with a recent finding indicating that a group of single nucleotide polymorphisms located in the PKG1 gene is associated with the salt‐sensitivity of blood pressure in hypertensive human subjects.28 Our previous studies indicate that marinobufagenin, acting via NKA‐dependent signaling, stimulates the synthesis of collagen in cardiac, renal, and aortic tissue13, 14, 15, 16, 19 and that immunoneutralization of marinobufagenin reverses cardiac fibrosis in the rat model of renal failure.29 Because fibrosis is a hallmark of cardiovascular aging,30, 31 the PKG agonists and natriuretic peptides exhibit antifibrotic effects.26, 32 The impact of the age‐associated reduction in cGMP/PKG signaling on the profibrotic effects of marinobufagenin merits further investigation.
Sources of Funding
This research was supported by the Intramural Research Program, National Institute of Aging, National Institutes of Health, Bethesda, MD (AG000609).
Disclosures
None.
References
| # | Year | Authors | Title |
|---|---|---|---|
| 1 | 1990 | Khaw KT , Barrett‐Connor E | Increasing sensitivity of blood pressure to dietary sodium and potassium with increasing age: a population study using casual urine specimens. Am J Hypertens. 1990;3:505–511. doi: 10.1093/ajh/3.6.505 2369500 [CrossRef] [PubMed] |
| 2 | 2001 | Sacks FM , Svetkey LP , Vollmer WM , Appel LJ , Bray GA , Harsha D , Obarzanek E , Conlin PR , Miller ER III , Simons‐Morton DG , et al | Effects on blood pressure of reduced dietary sodium and the Dietary Approaches to Stop Hypertension (DASH) diet. DASH‐Sodium Collaborative Research Group. N Engl J Med. 2001;344:3–10. doi: 10.1056/NEJM200101043440101 11136953 [CrossRef] [PubMed] |
| 3 | 2009 | Bagrov AY , Shapiro JI , Fedorova OV | Endogenous cardiotonic steroids: physiology, pharmacology, and novel therapeutic targets. Pharmacol Rev. 2009;61:9–38. doi: 10.1124/pr.108.000711 19325075PMC2763610 [CrossRef] [PubMed] |
| 4 | 2021 | Agalakova NI , Kolodkin NI , Adair CD , Trashkov AP , Bagrov AY | Preeclampsia: cardiotonic steroids, fibrosis, Fli1 and hint to carcinogenesis. Int J Mol Sci. 2021;22:1941. doi: 10.3390/ijms22041941 33669287PMC7920043 [CrossRef] [PubMed] |
| 5 | 1997 | Fedorova OV , Bagrov AY | Inhibition of Na/K ATPase from rat aorta by two Na/K pump inhibitors, ouabain and marinobufagenin: evidence of interaction with different alpha‐subunit isoforms. Am J Hypertens. 1997;10:929–935. doi: 10.1016/s0895-7061(97)00096-4 9270089 [CrossRef] [PubMed] |
| 6 | 2006 | Fedorova OV , Agalakova NI , Morrell CH , Lakatta EG , Bagrov AY | ANP differentially modulates marinobufagenin‐induced sodium pump inhibition in kidney and aorta. Hypertension. 2006;48:1160–1168. doi: 10.1161/01.HYP.0000248129.20524.d0 17043158 [CrossRef] [PubMed] |
| 7 | 2009 | Bagrov AY , Agalakova NI , Kashkin VA , Fedorova OV | Endogenous cardiotonic steroids and differential patterns of sodium pump inhibition in NaCl‐loaded salt‐sensitive and normotensive rats. Am J Hypertens. 2009;22:559–563. doi: 10.1038/ajh.2009.22 19229192PMC2678964 [CrossRef] [PubMed] |
| 8 | 2019 | Emelyanov IV , Konradi AO , Lakatta EG , Fedorova OV , Bagrov AY | Acute salt loading and cardiotonic steroids in resistant hypertension. Curr Top Membr. 2019;83:1–13. doi: 10.1016/bs.ctm.2019.01.005 31196601PMC7508670 [CrossRef] [PubMed] |
| 9 | 2015 | Fedorova OV , Lakatta EG , Bagrov AY , Melander O | Plasma level of the endogenous sodium pump ligand marinobufagenin is related to the salt‐sensitivity in men. J Hypertens. 2015;33:534–541; discussion 541. doi: 10.1097/HJH.0000000000000437 25479026PMC4543292 [CrossRef] [PubMed] |
| 10 | 2001 | Fedorova OV , Kolodkin NI , Agalakova NI , Lakatta EG , Bagrov AY | Marinobufagenin, an endogenous alpha‐1 sodium pump ligand, in hypertensive Dahl salt‐sensitive rats. Hypertension. 2001;37:462–466. doi: 10.1161/01.hyp.37.2.462 11230319 [CrossRef] [PubMed] |
| 11 | 2008 | Fedorova OV , Simbirtsev AS , Kolodkin NI , Kotov AY , Agalakova NI , Kashkin VA , Tapilskaya NI , Bzhelyansky A , Reznik VA , Frolova EV , et al | Monoclonal antibody to an endogenous bufadienolide, marinobufagenin, reverses preeclampsia‐induced Na/K‐ATPase inhibition and lowers blood pressure in NaCl‐sensitive hypertension. J Hypertens. 2008;26:2414–2425. doi: 10.1097/HJH.0b013e328312c86a 19008721PMC2583455 [CrossRef] [PubMed] |
| 12 | 2019 | Zhang Y , Wei W , Shilova V , Petrashevskaya NN , Zernetkina VI , Grigorova YN , Marshall CA , Fenner RC , Lehrmann E , Wood WH III , et al | Monoclonal antibody to marinobufagenin downregulates tgf β profibrotic signaling in left ventricle and kidney and reduces tissue remodeling in salt‐sensitive hypertension. J Am Heart Assoc. 2019;8:e012138. doi: 10.1161/JAHA.119.012138 31576777PMC6818028 [CrossRef] [PubMed] |
| 13 | 2012 | Fedorova OV , Kashkin VA , Zakharova IO , Lakatta EG , Bagrov AY | Age‐associated increase in salt sensitivity is accompanied by a shift in the atrial natriuretic peptide modulation of the effect of marinobufagenin on renal and vascular sodium pump. J Hypertens. 2012;30:1817–1826. doi: 10.1097/HJH.0b013e328356399b 22796708PMC3722363 [CrossRef] [PubMed] |
| 14 | 2007 | Elkareh J , Kennedy DJ , Yashaswi B , Vetteth S , Shidyak A , Kim EG , Smaili S , Periyasamy SM , Hariri IM , Fedorova L , et al | Marinobufagenin stimulates fibroblast collagen production and causes fibrosis in experimental uremic cardiomyopathy. Hypertension. 2007;49:215–224. doi: 10.1161/01.HYP.0000252409.36927.05 17145984 [CrossRef] [PubMed] |
| 15 | 2018 | Fedorova OV , Ishkaraeva VV , Grigorova YN , Reznik VA , Kolodkin NI , Zazerskaya IE , Zernetkina V , Agalakova NI , Tapilskaya NI , Adair CD , et al | Antibody to marinobufagenin reverses placenta‐induced fibrosis of umbilical arteries in preeclampsia. Int J Mol Sci. 2018;19:19. doi: 10.3390/ijms19082377 PMC612125630104471 [CrossRef] [PubMed] |
| 16 | 2014 | Haller ST , Drummond CA , Yan Y , Liu J , Tian J , Malhotra D , Shapiro JI | Passive immunization against marinobufagenin attenuates renal fibrosis and improves renal function in experimental renal disease. Am J Hypertens. 2014;27:603–609. doi: 10.1093/ajh/hpt169 24014658PMC3958603 [CrossRef] [PubMed] |
| 17 | 2010 | Trojanowska M | Cellular and molecular aspects of vascular dysfunction in systemic sclerosis. Nat Rev Rheumatol. 2010;6:453–460. doi: 10.1038/nrrheum.2010.102 20585340PMC3824624 [CrossRef] [PubMed] |
| 18 | 2015 | Fedorova OV , Emelianov IV , Bagrov KA , Grigorova YN , Wei W , Juhasz O , Frolova EV , Marshall CA , Lakatta EG , Konradi AO , et al | Marinobufagenin‐induced vascular fibrosis is a likely target for mineralocorticoid antagonists. J Hypertens. 2015;33:1602–1610. doi: 10.1097/HJH.0000000000000591 26136067PMC4547457 [CrossRef] [PubMed] |
| 19 | 2018 | Grigorova YN , Wei W , Petrashevskaya N , Zernetkina V , Juhasz O , Fenner R , Gilbert C , Lakatta EG , Shapiro JI , Bagrov AY , et al | Dietary sodium restriction reduces arterial stiffness, vascular TGF‐β‐dependent fibrosis and marinobufagenin in young normotensive rats. Int J Mol Sci. 2018;19:19. doi: 10.3390/ijms19103168 PMC621409330326586 [CrossRef] [PubMed] |
| 20 | 1997 | Pauly RR , Bilato C , Cheng L , Monticone R , Crow MT | Vascular smooth muscle cell cultures. Methods Cell Biol. 1997;52:133–154. doi: 10.1016/s0091-679x(08)60377-5 9379948 [CrossRef] [PubMed] |
| 21 | 2013 | Jablonski KL , Fedorova OV , Racine ML , Geolfos CJ , Gates PE , Chonchol M , Fleenor BS , Lakatta EG , Bagrov AY , Seals DR | Dietary sodium restriction and association with urinary marinobufagenin, blood pressure, and aortic stiffness. Clin J Am Soc Nephrol. 2013;8:1952–1959. doi: 10.2215/CJN.00900113 23929930PMC3817896 [CrossRef] [PubMed] |
| 22 | 2017 | Takahashi T , Asano Y , Sugawara K , Yamashita T , Nakamura K , Saigusa R , Ichimura Y , Toyama T , Taniguchi T , Akamata K , et al | Epithelial Fli1 deficiency drives systemic autoimmunity and fibrosis: possible roles in scleroderma. J Exp Med. 2017;214:1129–1151. doi: 10.1084/jem.20160247 28232470PMC5379967 [CrossRef] [PubMed] |
| 23 | 2009 | Elkareh J , Periyasamy SM , Shidyak A , Vetteth S , Schroeder J , Raju V , Hariri IM , El‐Okdi N , Gupta S , Fedorova L , et al | Marinobufagenin induces increases in procollagen expression in a process involving protein kinase C and Fli‐1: implications for uremic cardiomyopathy. Am J Physiol Renal Physiol. 2009;296:F1219–F1226. doi: 10.1152/ajprenal.90710.2008 19261738PMC2681369 [CrossRef] [PubMed] |
| 24 | 2022 | Grigorova YN , Juhasz O , Long JM , Zernetkina VI , Hall ML , Wei W , Morrell CH , Petrashevskaya N , Morrow A , LaNasa KH , et al | Effect of cardiotonic steroid marinobufagenin on vascular remodeling and cognitive impairment in young Dahl‐S rats. Int J Mol Sci. 2022;23:4563. doi: 10.3390/ijms23094563 35562955PMC9101263 [CrossRef] [PubMed] |
| 25 | 2022 | Numata G , Takimoto E | Cyclic GMP and PKG signaling in heart failure. Front Pharmacol. 2022;13:792798. doi: 10.3389/fphar.2022.792798 35479330PMC9036358 [CrossRef] [PubMed] |
| 26 | 2021 | Nakamura T , Tsujita K | Current trends and future perspectives for heart failure treatment leveraging cGMP modifiers and the practical effector PKG. J Cardiol. 2021;78:261–268. doi: 10.1016/j.jjcc.2021.03.004 33814252 [CrossRef] [PubMed] |
| 27 | 2022 | Caporarello N , Lee J , Pham TX , Jones DL , Guan J , Link PA , Meridew JA , Marden G , Yamashita T , Osborne CA , et al | Dysfunctional ERG signaling drives pulmonary vascular aging and persistent fibrosis. Nat Commun. 2022;13:4170. doi: 10.1038/s41467-022-31890-4 35879310PMC9314350 [CrossRef] [PubMed] |
| 28 | 2011 | Citterio L , Simonini M , Zagato L , Salvi E , Delli Carpini S , Lanzani C , Messaggio E , Casamassima N , Frau F , D’Avila F , et al | Genes involved in vasoconstriction and vasodilation system affect salt‐sensitive hypertension. PLoS One. 2011;6:e19620. doi: 10.1371/journal.pone.0019620 21573014PMC3090407 [CrossRef] [PubMed] |
| 29 | 2012 | Haller ST , Kennedy DJ , Shidyak A , Budny GV , Malhotra D , Fedorova OV , Shapiro JI , Bagrov AY | Monoclonal antibody against marinobufagenin reverses cardiac fibrosis in rats with chronic renal failure. Am J Hypertens. 2012;25:690–696. doi: 10.1038/ajh.2012.17 22378033PMC3355226 [CrossRef] [PubMed] |
| 30 | 2022 | Kobayashi M , Girerd N , Ferreira JP , Kevin D , Huttin O , Gonzalez A , Bozec E , Clark AL , Cosmi F , Cuthbert J , et al | The association between markers of type I collagen synthesis and echocardiographic response to spironolactone in patients at risk of heart failure: findings from the HOMAGE trial. Eur J Heart Fail. 2022;24:1559–1568. doi: 10.1002/ejhf.2579 35703355 [CrossRef] [PubMed] |
| 31 | 2005 | Scavone C , Munhoz CD , Kawamoto EM , Glezer I , de Sa Lima L , Marcourakis T , Markus RP | Age‐related changes in cyclic GMP and PKG‐stimulated cerebellar Na,K‐ATPase activity. Neurobiol Aging. 2005;26:907–916. doi: 10.1016/j.neurobiolaging.2004.08.013 15718050 [CrossRef] [PubMed] |
| 32 | 2018 | Hofmann F | A concise discussion of the regulatory role of cGMP kinase I in cardiac physiology and pathology. Basic Res Cardiol. 2018;113:31. doi: 10.1007/s00395-018-0690-1 29934662 [CrossRef] [PubMed] |